| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
Biomembrane celulare
Biomembranele reprezinta circa 10 - 20% din masa celulara. Proportia de lipide si proteine care intra in componenta acestora este diferita in raport cu functiile particulare ale fiecarei biomembrane. Membranele cu activitate metabolica marita contin mai multe proteine (de ex. membrana interna a mitocondriei cu peste 70% proteine in alcatuirea sa). Teaca mielinica contine lipide in proportie de 75% ceea ce asigura rolul sau de izolant electric. Glucidele reprezinta un component minor, prezent numai in proportie de 2-10%. Acestea sunt dispuse in principal pe fata externa a membranei fiind legate covalent de lipide sau proteine. Acestor componente la nivel membranar li se asociaza o insemnata cantitate de apa (circa 20% din masa totala) precum si ioni de Ca si Mg. Proportiile lor variaza foarte mult in special in stari patologice.
Proteinele membranare sunt unele periferice iar altele sunt integrate in lipide.
Pe fata citoplasmica a multor membrane celulare se formeaza o retea de proteine fibrilare formand citoscheletul.
Pe fata externa a membranelor atat proteinele cat si lipidele prezinta portiuni oligozaharidice fiind constituite ca glicoproteine si glicolipide.
Acestea formeaza in ansamblul lor invelisul celular numit glicocalix.
Lipidele membranare pot fi nepolare (neutre) cum este colesterolul ori polare cum sunt glicerofosfolipidele si sfingolipidele.
Modelul de membrana acceptat este cu dublu strat lipidic, monomoleculare juxtapuse cu capetele polare spre exterior si cu catenele hidrocarburice in contact.
Stratul lipidic este strabatut
de
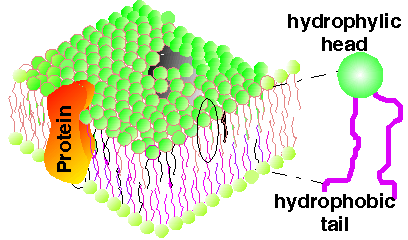
Modelul este de mozaic fluid de proteine globulare in dublul strat lipidic.
I. Stratul dublu reprezinta matricea lipidica a biomembranelor iar structura sa este realizata printr-un proces de auto-asamblare (autoorganizare).
Fortele de atractie Van der Walls dintre lanturile hidrocarbonate determina, la randul lor, aglomerarea acestora. De asemenea intre constituientii membranari se manifesta si interactiunile de tip electrostatic sau de tip legatura de H (intre capetele polare si moleculele de apa).
Proprietatea de autoorganizare a stratului dublu se pastreaza si in vitro, putandu-se astfel obtine membranele artificiale. Acestea se prezinta de obicei sub doua forme: fie ca straturi bi-lipidice plane, fie ca straturi duble lipidice dispuse in vezicule (lipozomi).
II. Fluiditate: Stratul dublu lipidic efectueaza anumite miscari intra sau intermoleculare, fiind inzestrat, prin insasi structura sa, cu o mare mobilitate.
Frecventa si amplitudinea fiecarui tip de deplasare a stratului dublu depind de caracteristicile sale mecanice si structurale. Astfel, miscarea cea mai simpla de observat o reprezinta rotatia lipidelor in jurul propriei axe moleculare si a lanturilor hidrocarbonate. Cu cat aceste miscari sunt mai ample cu atat mai insemnate devin si miscarile laterale ale lipidelor si proteinelor in planul membranei.
Moleculele lipidice prezinta si proprietatea de a difuza liber in stratul dublu lipidic.
~n afara acestor deplasari se mai evidentiaza si deplasari intramoleculare (in care se deplaseaza doar o parte a moleculei) miscari de rotatie de translatie a moleculei.
Miscarile de translatie ale moleculelor lipidice pot fi laterale sau de migrare dintr-un strat in altul.
Prin translatia laterala, moleculele lipidice isi schimba locul cu moleculele vecine in cadrul aceluiasi monostrat lipidic. Viteza acestei miscari este de circa 107 m/s. Migrarea lipidelor de la un strat la altul este mult mai redusa. Prin mecanismul basculant numit "flip-flop", anumite enzime realizeaza un transfer specific al unor fosfolipide dintr-un monostrat in celalalt. Aceasta se datoreste si compozitiei lipidice diferite a unui monostrat fata de monostratul vecin. Miscarea "flip-flop" este foarte lenta ceea ce face sa explice formarea dublului strat asimetric cu aparitie in cursul biosintezei acestuia in reticulul endoplasmatic.
Proteine membranare
Proteinele membranare sunt specifice fiecarei membrane. Ele reprezinta elementul activ al membranei (cu rol de enzime, canale, transportori, receptori, etc.).
Concentratia lor variaza functie de tipul membranei intre 20% si 75% (membrana mitocondriilor).
Putem grupa aceste proteine in doua categorii:
a) proteine extrinseci sau periferice, sau care patrund in membrana pe o anumita adancime. Ele sunt slab legate de membrana si pot fi indepartate prin spalare ori prin tratare cu solutie ionica scazuta. Ele sunt frecvent implicate in transmiterea informatiei spre interiorul celulei.
b) proteine intrinseci sau integrale care traverseaza membrana pe toata grosimea ei. Pot fi extrase prin tratare cu detergenti sunt implicate in general in fenomene de transport. Partile intramembranare sunt structuri hidrofobe, in dublu helix, legate intre ele, in mediul apos, prin zone hidrofile neelicoidale.
Moduri de legare a proteinelor membranare
proteine intrinseci
a) cu un singur segment transmembranar;
b) segmente transmembranare multiple.
proteine extrinseci
c) legaturi electrostatice;
d) legaturi hidrofobe;
e) legaturi hidrofobe, prin ancorare de un segment terminal ce penetreaza membrana;
f) ancorare printr-o molecula lipidica legata covalent;
g) legare de o alta proteina membranara.
Posibilitati de miscare in stratul bilipidic
Lipide
a) difuzie laterala in stratul lipidic (stanga) si schimb intre moleculele lipidice din strat si cele adiacente proteinelor (dreapta);
b) rotatie in jurul axei proprii si basculare dintr-un strat in celalalt;
c) rotatie descriind o suprafata conica (o asemenea miscare pot avea si derivatii acizilor grasi);
Proteine
d) difuzie laterala;
e) rotatie in jurul unei axe perpendiculare pe stratul bilipidic;
f) rotatie izotropica (fara axa preferentiala);
g) rotatie descriind suprafete conice, molecule hidrofobe, comportandu-se ca un bastonas rigid.
Transportul de substanta prin membrana celulara
Clasificarea mecanismelor de transport membranar
Transport pasiv (fara consum de energie metabolica, datorita gradientilor de concentratie, de potential electric, de presiune, gravitational si in sensul lor.
difuzie simpla prin stratul bilipidic;
difuzie prin canale;
difuzie facilitata de moleculele transportoare;
osmoza
Transport activ (consum de energie metabolica impotriva gradientilor, realizat de molecule transportoare, cu proprietati enzimatice.
primar (pompe ionice) este cuplat cu o reactie exergonica (exemplu hidroliza ATP);
secundar: este cuplat cu transportul simultan al unei alte substante in sensul gradientului electrochimic;
simport: ambele substante sunt transportate in acelasi sens;
antiport: cele doua substante sunt transportate in sensuri opuse.
Permeabilitatea si selectivitatea membranelor biologice
~n cazul organismelor vii intalnim membrane permeabile si selectiv permeabile constand in bariere de complexitati diferite:
bariere multicelulare (intestinul);
monocelulare (peretele vaselor capilare);
membranele particulelor subcelulare (nuclee, mitocondrii, lipozomi).
Membranele biologice au permeabilitati foarte mari pentru moleculele de apa fata de permeabilitatea pentru moleculele polare sau macromolecule.
Permeabilitatea este mare si pentru molecule sau pentru ioni hidratati.
Membranele pot fi clasificate dupa proprietatile lor de permeabilitate:
impermeabile;
permeabile;
selectiv permeabile:
q SMP:
naturale:
pluricelulare;
monocelulare;
celulare
artificiale:
organice;
anorganice
q IRP.
SMP - membrane semipermeabile (permeabile numai la solvent).
IRP - membrane ireciproc permeabile, adica permeabile la solviti dar numai intr-un sens.
Transportul pasiv
Este un proces de difuzie a ionilor si a moleculelor prin membrana.
~n acest transport pasiv exista trei fenomene distincte ca modalitate de transport microscopic.
difuzia libera prin membrana (dializa) sub actiunea unui gradient de concentratie;
filtrarea printr-o membrana sub actiunea unui gradient de presiune si concentratie.
Aceste fenomene se intampla pana cand sistemul ajunge in echilibru termodinamic avand aceleasi valori de concentratie si presiune intre mediul intra si extracelular.
Difuzia simpla
Datorita structurii membranei ca strat bilipidic, zona interna fiind hidrofoba o particula trebuie sa strabata o zona hidrofila si apoi sa ajunga in zona hidrofoba. Datorita acestui lucru mecanismele de difuzie vor fi diferite pentru particulele hidrofile (ioni si molecule polare) si particule hidrofobe (nepolare) respectiv pentru particulele liposolubile si hidrosolubile.
Solubilitatea unei substante este diferita in mediul apos si in membrana.
Raportul intre solubilitatea intr-un solvent (in cazul nostru membrana lipidica fluida) si solubilitatea in apa se numeste coeficient de partitie solvent-apa (b
Un coeficient de partitie diferit de 1 determina un salt de concentratie pe suprafata membranei.
Difuzia pasiva sau transportul pasiv datorat agitatiei termice a unor particule din zonele de concentratie (densitate mai ridicata) spre zonele cu valori mai mici printr-un mediu suport omogen se supune la doua legi cunoscute sub numele de legile lui Fick.
Pentru prezentarea legilor difuziei este necesar a utiliza doi termeni: flux si gradient.
Fluxul reprezinta cantitatea de substanta, sarcina, energie etc., transportate printr-o suprafata S in unitate de timp.
Flux masic Dm/Dt (la limita (dm/dt)
Flux molar Dn Dt (la limita dn/dt).
Prin gradient se intelege variatia unei marimi (concentratie, densitate, potential electric etc.) intre doua puncte ale spatiului raportata la distanta dintre cele doua puncte.
Prima lege a lui Fick:
Fluxul masic dintr-o substanta printr-o suprafata S este direct proportionala cu suprafata si cu gradientul de concentratie:
Legea a II-a a lui Fick
Este o lege de conservare. ~n procesul difuziei in intervalul de timp (t, t + at) masa de solvit ce intra in volumul Sdx este egala cu masa ce ramane in acest volum, minus masa de lichid care iese,
Variatia temporala a concentratiei in orice punct al solutiei este proportionala cu variatia spatiala a gradientului de concentratie.
Aplicatii ale difuziei
q Transportul intracelular;
q Transporturile prin membrana sau dializa;
q Filtrarea printr-o membrana;
q Filtrarea unei solutii;
q Osmoza si presiunea osmotica.
Difuzia facilitata
Dupa cum s-a vazut, moleculele mici hidrofobe pot traversa membrana prin difuzie simpla, iar ionii difuzeaza prin canale.
Moleculele hidrofile mari (care constituie factori nutritivi pentru celule nu pot traversa membrana decat daca utilizeaza un transport mediat de o molecula transportoare, numita ca mecanism difuzie facilitata.
Molecula transportoare recunoaste molecula de transportat pe care o introduce in celula in sensul gradientului electrochimic.
Transportorii sunt capabili sa deosebeasca izomerul dextrogir de cel levogir.
Difuzia facilitata poate fi descrisa pe baza cineticii Michaelis - Menten.
Descrierea mecanismului:
Molecula transportoare, cu rol enzimatic, se poate afla in doua stari conformationale:
avand un loc de legare a substratului (molecula ce urmeaza a fi transportata) pe una din fetele membranei;
modificarea conformationala ca rezultat de transferare a situsului pe cealalta fata a membranei.
Viteza de reactie este proportionala cu concentratia substratului, transportul se efectueaza de la concentratie mare la concentratie mica (in directia gradientului de concentratie a substratului). Similar altor reactii enzimatice difuzia facilitata poate sa fie inhibata de anumiti compusi prin inhibitie competitiva sau necompetitiva.
Eficienta difuziei facilitate este mult mai mica decat a celei prin canale. Ex: transportul de glucoza din membrana eritrocitara mediaza circa 300 molec/s.
Un caz particular al difuziei facilitate este formarea in jurul ionului a unei structuri hidrofobe, care ii permite difuzia prin stratul bilipidic.
Difuzia prin canale
Substantele ionizate nefiind liposolubile, difuzia lor prin membrana se poate face prin structuri proteice specializate care strabat membrana pe toata grosimea ei si creeaza cai de trecere pentru ioni, formand canale (ionice).
Aceste canale permit trecerea ionilor in ambele sensuri
directionalitatea fiind impusa de gradientul electro-chimic. Notiunea veche era
de
Canalul ionic poate fi o enzima (E) care formeaza tranzitoriu un complex care va fi transportat ele formand substratul (S). Functionarea sa se face pe baza cineticii Michaelis-Menten.
Canalul se poate inchide si deschide functie de modificarea conformationala a proteinei canal. Comanda se poate realiza:
electric prin modificarea potentialului membranar (canale Na+ si K+);
chimic prin molecule ce se leaga specific la proteina canal;
prin alte mecanisme.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3075
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2026 . All rights reserved