| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Membrana plasmatica
Membrana plasmatica inconjura celula bacteriana si este bariera separatoare a citoplasmei de mediul extern. Consecinta imediata a lezarii membranei este pierderea componentelor citoplasmatice.
Grosimea membranei este de 7-10 nm. La microscopul electronic are o structura trilaminara, dupa modelul unitar al membranelor celulare (unit membrane) al lui Robertson. Pe baza structurii fine s-a considerat (eronat) ca membrana plasmatica este alcatuita din doua straturi de proteine, intre care se gaseste unul lipidic, dar cantitatea de proteine este prea mica pentru a forma doua straturi continui.
Membrana celulei procariote contine proteine, glucide si lipide, dar spre deosebire de membrana celulei eucariote nu contine steroli (fig. 15a).
Singer si Nicolson au propus modelul mozaicului fluid (fig. 15b) de organizare a membranelor biologice, in acord cu care membranele sunt structuri bidimensionale de proteine globulare si lipide, cu distributie orientata, stabilizate de cea de a III-a componenta majora - apa. Moleculele de apa sunt legate prin punti intermoleculare de H, formand o structura de retea. Dizolvarea unei molecule in apa semnifica stabilirea unei continuitati intre structura chimica a moleculei dizolvate si structura de retea a apei. Pentru a se integra (dizolva) in structura de retea de apei, molecula trebuie sa formeze o legatura de H cu apa sau sa accepte o legatura a acesteia.
Fig. 15. a. Reprezentarea schematica a moleculei fosfolipidice, componenta structurala a membranei. Fosfolipidele sunt molecule polare. "Cozile" de acizi grasi sunt foarte hidrofobe (nu formeaza legaturi cu apa) si constituie o bariera de permeabilitate fata de moleculele hidrosolubile. "Capul" moleculei este format din gruparea fosfat, legata de o grupare care contine N si din glicerol. Este foarte hidrofil (formeaza legaturi cu moleculele de apa).
b. Modelul mozaicului fluid al structurii membranei. Fosfolipidele formeaza un strat dublu, cu componentele hidrofobe orientate spre interior, iar capetele hidrofile constituie suprafata interna si externa a membranei. In "marea" lipidica proteinele plutesc ca niste "iceberg-uri". Unele se extind in toata grosimea dublului strat lipidic, iar altele sunt ancorate pe fata interna sau externa. Membrana micoplasmelor si eucariotelor contine colesterol.
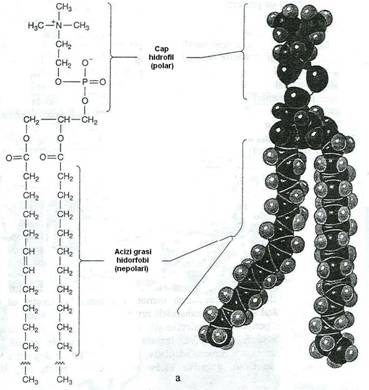

Membrana este un dublu strat fosfolipidic si glicolipidic, la care se asociaza proteinele membranare. Ansamblul molecular al membranei este asemanat cu un ocean fosfolipidic, in care plutesc ca niste iceberguri, proteinele.
Proteinele confera membranelor, multe dintre proprietatile functionale specifice (de exemplu, mentinerea si utilizarea gradientului transmembranar de H pentru sinteza ATP).
In raport cu dispunerea lor in structura membranei, proteinele sunt periferice si integrate.
Proteinele periferice* sunt asociate cu membrana, dar nu au nici o secventa inclusa in structura ei. Structura lor este analoga proteinelor hidrosolubile.
Clasa proteinelor periferice este impartita in doua subclase:
proteinele asociate membranei sunt globulare, legate pe suprafata membranei, fie de stratul lipidic, fie de proteinele integrate;
proteinele membranare de schelet formeaza scheletul membranei si sunt localizate imediat sub membrana plasmatica a celulei. Ele se asociaza componentelor membranei, iar in celula eucariota se leaga si cu citoscheletul. La procariote se accepta existenta unui citoschelet primitiv, format din EF-Tu Elongation Factor Termo-unstable) si FtsZ (Factorul termo-stabil), cu rolul de suport al structurii celulare. Lipsesc structurile de tipul microfilamentelor si microtubulilor. FtsZ are potential de polimerizare si are functia se retea celulara de suport. Echivalentul lui EF-Tu in celula eucariota este EF1a.
Proteinele membranare integrate au cel putin un domeniu al moleculei situat in regiunea hidrofoba a stratului lipidic. Ele pot fi dislocate din structura membranei numai sub actiunea detergentilor ce solubilizeaza lipidele.
Clasa proteinelor integrate se imparte in doua subclase:
Proteinele transmembranare au o mare parte a masei lor inclusa in dublul strat fosfolipidic, dar expun domenii semnificativ diferite pe ambele fete ale membranei, ceea ce confera asimetria functionala a acesteia.
Ele au o structura heterogena, deoarece cel putin un domeniu al moleculei este inclus in mediul hidrofob al lipidelor si trebuie sa fie compatibil cu acesta. Secventa de 19-23 aminoacizi a domeniului inclus in stratul lipidic este hidrofoba si are configuratia a-helix (fig. 17).
Domeniile extralipidice ale proteinelor integrate sunt hidrofile, ceea ce confera asimetria structurala si functionala a membranei. Proteinele integrate difuzeaza liber in planul lateral al matricei lipidice, dar nu trec liber dintr-un strat in altul si de aceea asimetria functionala a membranei se pastreaza pentru perioade lungi. Asa se explica permeabilitatea superioara a fetei interne in raport cu fata externa.
Asimetria structurala a proteinelor integrate se evidentiaza pe imagini electrono-optice ale membranei criofracturate (fig. 16). Tehnica criofracturarii presupune inghetarea rapida a membranei la temperatura azotului lichid si fracturarea membranei cu un cutit special. Membrana se cliveaza de-a lungul regiunii hidrofobe a acizilor grasi si rezulta doua jumatati de membrana, cu un grad accentuat de asimetrie, conferita de proteinele transmembranare.
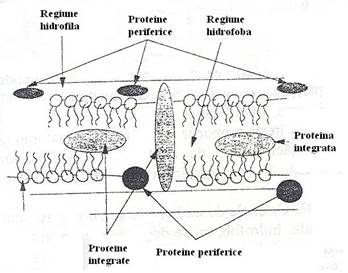
Fig. 16. Reprezentarea schematica a localizarii proteinelor membranare. Proteinele periferice sunt in raporturi spatiale stranse cu capetele hidrofile ale fosfolipidelor. Proteinele integrate sunt localizate in dublul strat fosfolipidic ori au domenii care se extind in regiunile hidrofile (dupa Holt si colab., 1998).
2. Proteinele membranare ancorate au un domeniu ce penetreaza dublul strat lipidic, dar nu traverseaza complet membrana. Sunt ancorate de membrana prin legatura covalenta cu lipidele.
Structura membranei plasmatice este stabilizata prin legaturi de H si interactiuni moleculare hidrofobe. Ionii de Mg si Ca realizeaza legaturi ionice cu sarcinile negative ale fosfolipidelor si stabilizeaza structura membranei.
Lipidele contin esteri ai acizilor grasi cu glicerolul. Predomina acizii cu l4-l6 atomi de C, saturati sau nesaturati.
Fosfolipidele (fosfogliceride) membranelor biologice sunt molecule amfipatice polare, deoarece au un cap polar (glicerolul) legat cu cozile nepolare ale acizilor grasi: glicerolul intra in structura de retea a apei, fiind hidrofil, iar acizii grasi formeaza componenta hidrofoba a moleculei (fig. 17). Astfel, membrana are doua suprafete polare, hidrofile, datorita resturilor de glicerol.
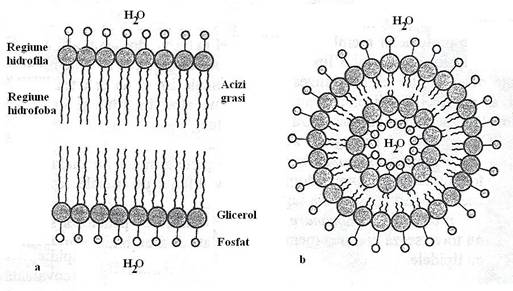
Fig. 17. a. Structura de baza a dublului strat fosfolipidic. b. Distributia stabila a moleculelor lipidice in apa, cu formarea unei vezicule membranare.
Intr-o solutie apoasa, lipidele se agrega si formeaza spontan structuri in dublu strat: acizii grasi la interior, in mediul hidrofob, iar moleculele de glicerol raman expuse in mediul apos. Agregatul functioneaza ca o bariera impermeabila pentru trecerea libera a moleculelor polare in si din celula.
Dublul strat lipidic este o structura ordonata. Catenele de carbon ale acizilor grasi sunt constranse la o asezare paralela si datorita caracterului hidrofob, mobilitatea lor in afara stratului lipidic este foarte limitata. Moleculele fosfolipidice sunt foarte mobile si se deplaseaza in plan lateral (bidimensional), in acelasi strat, cu o frecventa foarte mare, dar deplasarile dintr-un strat in celalalt (tridimensionale) sunt foarte rare (una la cateva ore).
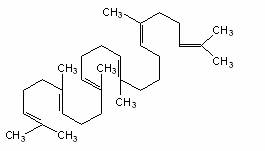
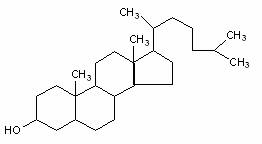
Squalen Colesterol
Structura moleculara a squalenului (9cu 30 atomi de C), precursorul biosintetic al sterolilor si structura colesterolului (un sterol tipic).
Rolul lipidelor membranare, s-a studiat folosind ca model, veziculele membranare artificiale sau naturale, ce se formeaza spontan dupa spargerea celulelor. S-a demonstrat astfel ca procariotele isi regleaza fluiditatea membranei, prin modificarea proportiei acizilor grasi saturati/nesaturati. Cultivarea la temperaturi scazute mareste proportia acizilor grasi nesaturati si sporeste gradul de fluiditate a membranei. Prin racire, faza de cristal lichid a lipidelor membranare trece spre o stare solida de gel, situatie in care grosimea dublului strat creste datorita extensiei catenelor de C ale lipidelor.
Stratul lipidic este suportul proteinelor si are rolul unei bariere de permeabilitate. Permeabilitatea diferentiata a celor doua fete ale membranei se explica atat prin asimetria proteinelor, cat si prin compozitia chimica diferita a lipidelor in cele doua straturi ale membranei.
Cu exceptia micoplasmelor si a metanotrofelor, membrana procariotelor se deosebeste de cea a eucariotelor prin absenta sterolilor. Sterolii sunt molecule plane, rigide, iar acizii grasi sunt flexibili. Sterolii confera un grad superior de rigiditate membranei plasmatice. Rigiditatea membranei este necesara celulelor lipsite de perete. La eucariote, rigiditatea ar fi necesara pentru a suporta fortele fizice care se exercita asupra membranei.
Antibioticele polienice (nystatin, candicidin) reactioneaza cu sterolii si destabilizeaza membrana. De aceea ele sunt active fata de celulele eucariote si nu ifluenteaza celulele procariote.
Micoplasmele incorporeaza in structura membranei, sterolul disponibil in mediul de crestere si sunt sensibile la antibioticele polienice.
La unele bacterii, in structura membranei se gasesc molecule asemanatoare structural cu colesterolul si pot avea acelasi rol de crestere a rigiditatii: hopanoidele. Un compus cu distributie larga este diploptenul, cu 30 atomi de C
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3834
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2026 . All rights reserved