| CATEGORII DOCUMENTE |
| Demografie | Ecologie mediu | Geologie | Hidrologie | Meteorologie |
Schimbari si modificari in biocenoze. Evolutia biocenozei
Precizare. Am abordat la acest nivel evolutia biocenozei deoarece modificarile si schimbarile care au loc sunt evidente la nivelul viului, al florei, microorganismelor si animalelor (Brewer, 1988). Se intelege ca in realitate este vorba despre o dinamica a ecosistemelor, deoarece odata cu biocenoza se schimba si se modifica si biotopul si factorii de mediu.
Bioceneza are o caraceteristica fundamentala si specifica viului: este dinamica si se schimba constant. Un copac moare, altul ii ia locul. O padure si implicit animalele din ea, se schimba necontenit: una este fenologia iarna, alta primavara, alta vara si alta toamna (Fig. 6-53). In acest caz este vorba despre o ritmicitate sezoniera. Pe linga acestea exista si alte tipuri de schimbari.
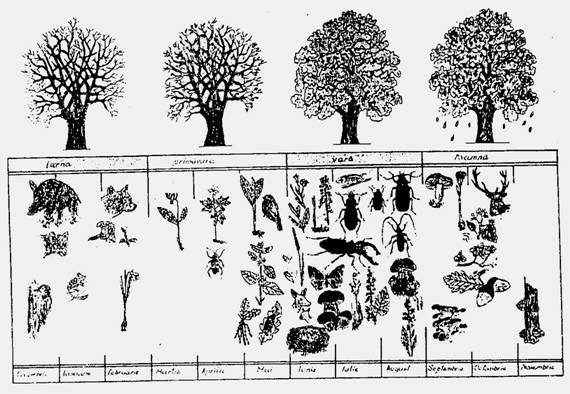
Fig. 6-53. Aspecte sugestive pentru ilustrarea schimbarilor la nivelul biocenozelor (ritmicitatea sezoniera) (dupa Fabian si Onaca, 1999)
1. Schimbari si modificari nedirectionate si directionate
Biocenoza (si implicit ecosistemul) sunt supuse schimbarilor. O prima categorie grupeaza schimbarile in nedirectionate si directionate (Brewer, 1988):
A. ☼ - schimbari nedirectionate (schimbari care nu afecteaza permanent biocenoza). Sunt incluse aici:
☻ - modificarile diurne si nocturne (vezi Fig. 6-29; lanturi trofice diurne si nocturne)
☻ - modificarile sezoniere (vezi Fig. 6-53).
☻ - schimbari de inlocuire (modificari aleatorii). Acestea contribuie la mentinerea stabilitatii biocenozei (ex. locul unui pradator este luat de catre altul; o veriga dintr-un lant trofic se schimba cu alta etc);
☻ - fluctuatiile (fluctuatiile biocenozelor sunt asociate cu fluctuatiile factorilor fizici de mediu).
B. ☼ - schimbarile directionate (sunt modificari inregistrate in structura biocenozelor, datorita modificarilor pe termen lung a factorilor de mediu; aceste schimbari au influente multiple asupra biocenozei. In aceasta categorie intra:
☻ - extinctia (disparitia speciilor dupa fenomene de schimbare majora a factorilor de mediu (vulcanism, cutremure, glaciatie);
☻ - succesiunile = ecogeneza (un proces de dezvoltare a unei biocenoze ce tinde sa atinga un stadiu de stabilitate).
2. Schimbari si modificari aritmice
Alti autori saugereaza alte clasificari ale schimbarilor de la nivelul biocenozelor si ecosistemelor Acestea sunt clasificate in: aritmice, ritmice si succesiuni (Stugren, 1965, 1975, 1994).
Schimbarile si modificarile aritmice (accidentale), nu au un caracter ciclic impus de factorii abiotici ai mediului iar biota este modificata asociat cu modificarile zilnice ale factorilor de mediu (ex. daca ploua unele insecte si rozatoare sunt inactive; daca este soare buprestidele sunt active; cand soarele intra in nori, albinele inceteaza zborul; o ploaie torentiala sau incendiu pot modifica distructiv biota unui spatiu etc) (cf. clasificarii anterioare, aici intra fluctuatiile).
3. Schimbari si modificari ritmice
Schimbarile si modificarile ritmice se desfasoara mai mult sau mai putin periodic, fiind determinate de variatiile periodice ale factorilor de mediu si implicit de ritmicitatea materiei vii (protoplasama functioneaza ritmic; deci sistemele vii functioneaza ritmic, ele facand parte dintr-un univers ritmic - Stugren, 1975). Aceste modificari sunt specifice sistemelor care functioneaza dupa schema feed back, mecanism ce determina oscilatii permanente in sistemul respectiv. Asemenea ritmuri sunt deci corelate cu ritmicitatea factorilor meteorologici si cosmici. Ritmul biologic este un ritm intern, fixat genetic, dar intotdeauna acesta se ajusteaza dupa ritmul exogen (al mediului inconjurator) si rezulta astfel ritmurile ecologice.
In aceasta categorie sunt incluse: ritmul diurn si ritmul sezonier.
☻ - Ritmul diurn (nictemeral, circadian) [lat. diurnus = zi] [gr. nyx, nyktos = noapte; hemera = zi] [lat. circa = aproximativ, cam; diem = zi]. Toate organismele, deci biocenozele, functioneaza in conditiile de alternanta a fazei de lumina cu cea de intuneric, a alternantei dintre zi si noapte. Intr-una din faze este activa o parte a biocenozei, iar in cealalta, o alta parte. In sistemele acvatice o caracteristica a ritmului circadian este migratia pe verticala a planctonului. In ecosistemele terestre acest ritm se manifesta prin operatiile de inlocuire a activitatilor speciilor in aceeasi nisa ecologica (organismele componente fac parte din doua categorii: diurne si nocturne - vezi Fig. 6-29) si implicit prin modificarea functiilor vitale (curba transpiratiei la plante, cantatul la animale etc) (Fig. 6-54).
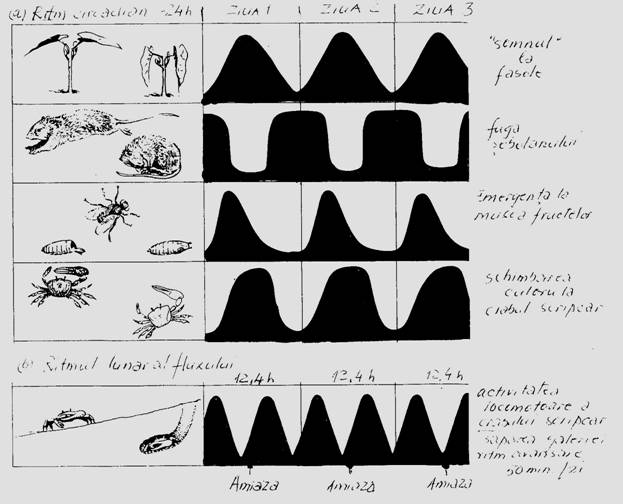
Fig. 6-54. Evidentierea unor modele comportamentale care ilustreaza activitatea ritmica circadiana, evidentiata in conditii experimentale (dupa Brown, 1959; din Smith, 1974).
Dupa cum am prezentat in cap. referitor la factorii fizici de mediu si organismul, se intelege ca si biocenoza reactioneaza la fotoperiodism (alternanta dintre fotofaza si scotofaza) printr-un "fotoperiodism de activitate" (plantele inceteaza noaptea fotosinteza, elimina CO2, unele specii de animale dorm, altele sunt active etc). In ecosistemele terestre ritmicitatea endogena a vegetatiei este mai pronuntata in stratul ierbos. In ceea ce priveste entomofauna, ritmul de activitate si densitatea carabidelor sunt determinate endogen in padure si pasuni. Pe solul padurii coleopterele pradatoare sunt active diferentiat: unele specii noaptea iar altele ziua; in timpul verii sunt active numai ziua. In padurea tropicala, insectele fitofage au activitatea si densitatea reduse in fazele de lumina tocmai pentru a evita pradatorii. La insectele nocturne la tropice s-a evidentiat si o periodicitate ca raspuns la lumina lunii dar in Europa activitatea insectelor este diminuata la lumina lunii, dar ea nu inceteaza asa cum sustin unele lucrari (aut.cit.in Stugren, 1994).
Organisme marine si-au adaptat ritmul endogen bimodal la ritmul bimodal al mareelor (Palmer, 1973). Specia Dictyota dichotoma (alge brune) (Cloudsley-Thompson, 1961) sau Uria aalge (pasare marina) (Slater, 1976) au un ritm al activitatii corelat cu fazele ciclului lunar.
In cadrul biocenozelor, speciile de consumatori constituie unitati functionale temporale, conform celor 4 momente care marcheaza alternanta: ziua, noaptea, crepusculul de seara (amurg), crepuscul de dimineata (zori) (Fig. 6-55, a). In conditii experimentale de laborator acest lucru este foarte evident (Fig. 6-55, c).
☻ - Ritmul sezonier (annual). Este caracterizat de alternanta anotimpurilor (specific in zona temperata) situatie in care se modifica durata perioadei de lumina, temperatura, precipitatiile, curentii de aer etc. Schimbandu-se sezonier macroclima, aceasta se repercuteaza si asupra biocenozelor care de data aceasta se succed in unitati functionale sezoniere (Fig. 6-54, b). Apoi, in fiecare sezon, biocenoza are o componenta activa si una pasiva care se inlocuiesc sezonier (Tischler, 1955).
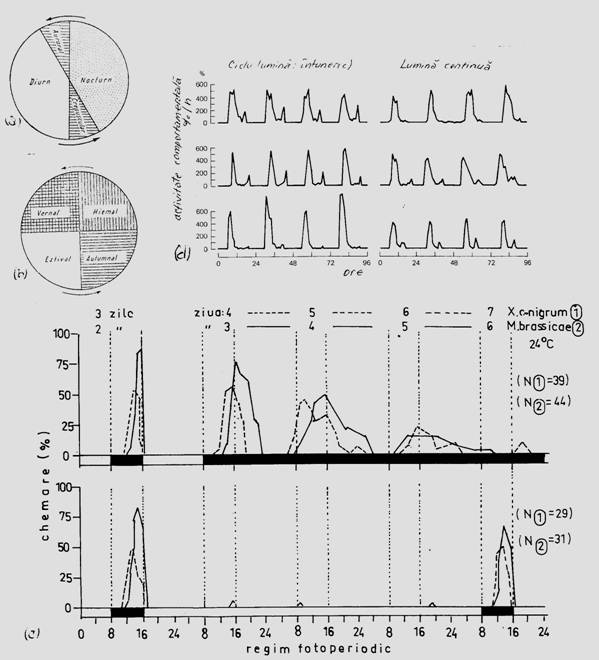
Fig. 6-55. Reprezentarea schematica a digramei ritmului diurn (a) si sezonier (b) al biocenozei. Evidentierea ritmului cricadian al comportamentului de chemare la femelele de Mamestra brassicae si Xestia c-nigrum (Lepidoptera: Noctuidae) (comportament nocturn) in conditii experimentale de laborator. Dupa 3 zile in care s-a pastrat ritmul de 16:8 ore (lumina:intuneric) in conditii de iluminare sau intuneric continuu in urmatoarele 3 zile, ritmicitatea a fost afectata (c). Evidentierea ritmului circadian la specia Chloris chloris (comportament diurn) in 3 situatii experimentale in care alternanta zi-noapte (lumina:intuneric) s-a modificat. Dupa 4 zile experimentul a continuat la lumina continua. Se constata modificari ale ritmicitatii, dar fiind specie diurna, la lumina continua aceste modificari nu sunt semnificative (d) (dupa Allee si colab., 1949; din Stugren, 1965; Stan, 1996).
Anumite parti ale biocenozei sau anumite lanturi trofice sunt active numai in anumite parti din an (fiind inactive in altele), iar unele elemente componente se pot schimba prin inlocuire, iar alteori se modifica gradul de dominanta dintre specii. Vulpes vulpes (vulpea roscata) si Ursus arctos (ursul brun) isi schimba ritmul de hranire sezonier in functie de periodicitatea activitatii plantelor si animalelor care constituie baza trofica a acestor specii (Fig. 6-56). Diferite parti structurale ale biocenozelor sau secvente din acestea au o ritmicitate sezoniera (ex. in padurile de fag curculionidele migreaza primavara spre coronamentul arborilor, iar toamna coboara in frunzar si in sol (Overgaard-Nielsen, 1974). In biotopuri aride colembolele formeaza doua componente sezoniere ce se inlocuiesc (Poinsot-Balanguer, 1976). Peste ritmul sezonier se sincronizeaza ritmul circadian.
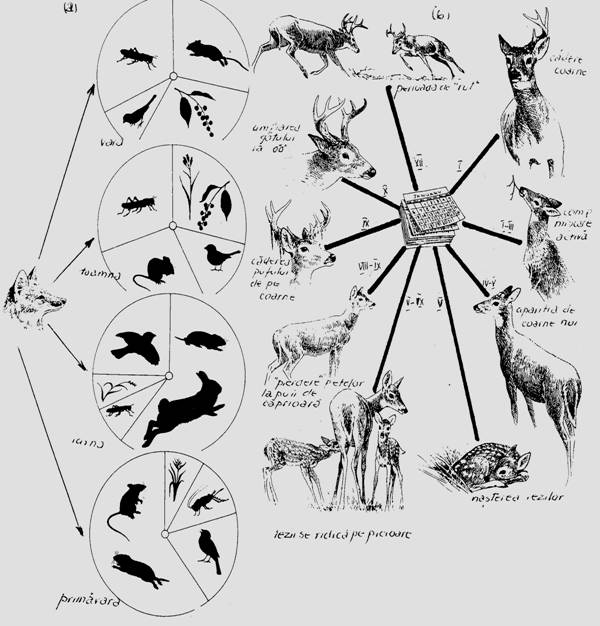
Fig. 6-56. Variatii la nivelul bazei trofice la vulpea roscata, dependent de periodicitatea sezoniera a timpului de dezvoltare a vegetatiei si activitatii animalelor care constituie sursa de hrana (a); activitatea comportamentala sezoniera la Cervus elaphus canadensis (b) (dupa Scott, 1955; din Smith, 1974).
Fitoplanctonul prezinta de asemenea, oscilatii ritmice sezoniere evidentiate prin variatia numarului de indivizi si a diversitatii specifice. Productivitatea in ocean inregistreaza oscilatii sezoniere (Zelikmann, 1977). Speciile animale marine prezinta modele comportamentale cu manifestare sezoniera si chiar si unele specii abisale (care se comporta aperiodic) manifesta ritmicitate reproductiva (Rokop, 1974).
Un comportament ritmic deosebit de interesant a fost studiat la anelidul Eunice viridis (Polichaeta) care si-a ajustat un ritm endogen al reproducerii, corelat cu o anumita faza a lunii (o zi inainte de ultimul patrar al lunii). Atunci indivizii se ridica la suprafata in numar mare iar partea posterioara a viermelui (hectocotilul), care reprezinta partea reproductiva, se desprinde de partea vegetativa si plutesc in cantitati mari, ceea ce implica si o ritmicitate a pradatorilor.
Pentru zona climatica temperata a Europei ritmul sezonier al biocenozelor este caracterizat de 6 aspecte caracteristice (configuratii):
☺ - hiemal (de iarna; pe perioada noiembrie - martie). Este perioada cind majoritatea poikilotermelor si homeotermelor sunt in repaos. Se manifesta activitate intensa la specii sedentare, comune, si la organismele din edafon si sub frunzar. Sunt functionale de asemenea si unele biocenoze aritmice (abisul oceanelor; adincul litosferei);
☺ - prevernal (de primavara timpurie; martie - aprilie). Este tranzitia de la iarna la vara cand fluxul energetic este slab, biomasa si metabolismul sunt reduse. Odata cu aparitia primelor forme de vegetatie incepe reactivarea fitofagilor si zoofagilor. Incep sa zboare primele insecte care au iernat ca adulti (lepidoptere, diptere, himenoptere etc), Se activeaza molustele si pedofauna. Incepe reconstituirea covorului ierbos dar cu specii putine, in schimb fauna devine activa;
☺ - vernal (de primavara;
aprilie - mai). Se intensifica biomasa si fluxul energetic. Se
compactizeaza stratul ierbos iar arborii au
☺ - estival (de vara; iunie - iulie). Activitatea la nivelul lanturilor trofice si retelelor trofice este maxima. Fluxul energetic si de substanta sunt maxime. Creste biomasa, metabolismul are valori mari. Se intensifica activitatea la toate nivelele dar se remarca ponderea uriasa a insectelor (Fig. 6-57).
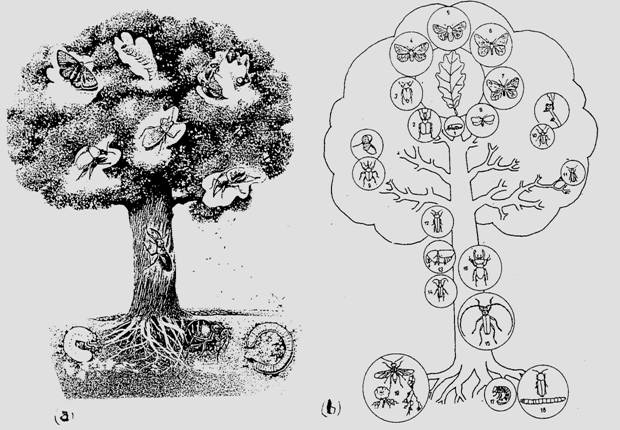
Fig. 6-57. Aspect sugestiv al predominantei entomofaunei (ilustrata aici in stratificare pe verticala) in configuratia estivala a ritmului sezonier. (a) aspect general; (b) detalii privind specii caracteristice ale entomofaunei (dupa Larousse, 1993; Fabian si Onaca, 1999). 1-8 consumatori ai frunzelor (1 - Orchestes quercus; 2 - Attelabus curculionides; 3 - Melolontha melolontha; 4 - Euproctis chryssorhoea; 5 - Malacosoma neustria; 6 - Eranis defoliaria; 7 - Operophtera brumata; 8 - Tortrix viridana); 9 consumator de fructe (Balaninus sp); 10 consumator de muguri (Phyllobius piri); 11-14 specii comune pe ramuri si scoarta arborilor; (11 - Cantharis obscura; 12 - Agrilus viridis; 13 - Scolytus intricatus; 14 - Rhagium sp.); 15-16 specii caracteristice pe tulpina; (15 - Cerambyx cerdo; 16 - Lucanus cervus); 17-19 specii endogee; (17 - larve de Melolontha melolontha; 18 - Agriotes lineatus; 19 - Cynips aptera).
☺ - serotinal (de toamna timpurie; august - septembrie). Structural biocenoza este asemanatoare cu cea din configuratia estivala. Vegetatia mai este relativ activa, fauna este activa si foarte activa, dar fluxul energetic se diminueaza, se reduce biomasa, structura functionala la nivelul lanturilor trofice si retelelor trofice se simplifica (pleaca pasarile migratoare; specii indigene intra in faze de latenta etc);
☺ - autumnal (toamna; septembrie - octombrie). Fluxul energetic, biomasa, metabolismul - se reduc la unele verigi. Vegetatia intra in faza de latenta dar raman foarte active organismele animale din frunzar si humus. Vietuitoarele mari isi diminueaza acivitatea locomotoare si hranirea, executa deplasari locale in vederea hibernarii.
Pe parcursul acestor configuratii, in ordinea prezentata mai sus, biocenoza evolueaza de la o compozitie saraca in specii si cu functionalitate redusa la minim, spre una bogata in specii si cu activitate complexa, mai ales in fazele serotinal - estival. Se subintelege ca aceasta ritmicitate caracterizeaza si domeniul subteran (Vandel, 1964).
4. Schimbari de inlocuire
In biocenoze stabile, organismele au tendinta de a avea o longevitate mare in acea comunitate. Aceasta se refera la afinitatea pentru acea comunitatea si capacitatea de a se mentine perioade de timp lungi ca veriga stabila in lanturile trofice constituite. Astfel sunt citate unele specii de ferigi cu o vechime de cca. 4000 de ani (Harlow si Harrar, 1968), iar unele specii de desert au 11.700 ani (Vasek, 1980). Dintre speciile de animalesunt amintite: crocodilii, dar si nevertebrate (ex. colembolele).
Exista insa in
cadrul majoritatii speciilor, unele care dupa anumite intervale
de timp, in urma modificarii radicale a conditiilor de mediu, dispar
din biocenoza si sunt inlocuite cu altele. Daca am vizitat o
padure matura acum 25 de ani si ne reintoarcem in ea in prezent,
aparent ea are aceeasi fizionomie si pare neschimbata. In
realitate aceasta
Un model clasic de un
asemenea ciclu la scara mica, dependent de caracteristicele plantelor
este un ciclu descris pentru mlastinile din
|
|
| ||||||
|
|
Movila de Sphagnum |
Calluna sp Juncus sp. Scirpus sp. |
|
||||
|
|
|
|
Calluna sp. Licheni |
||||
|
Portiuni de Sphagnum usor ridicate |
Calluna sp. moarta si terenul ridicat erodat; alte ridicaturi apar alaturi | ||||||
|
|
Balti mici |
|
In desertul Chihuahuan, doua importante specii Larrea tridentata si Opuntia leptocaulis sunt cuplate intr-un sistem ciclic de inlocuire, iar ciclul este partial condus de animalele din biocenoza (Yeaton, 1978) (Fig. 6-58). Larrea tridentata serveste ca suport si refugiu pentru pasari si mamifere care au mancat seminte si fructe de Opuntia. Ulterioe aceasta creste si se dezvolta sub Larrea, iar sistemul sau de radacini se dezvolta superficial si printre ramurile de Larrea. In final Larrea moare si se dezvoltra Opuntia. Actiunea vantului, rozatoarelor si a apei, expun radacinile plantei care in final moare.
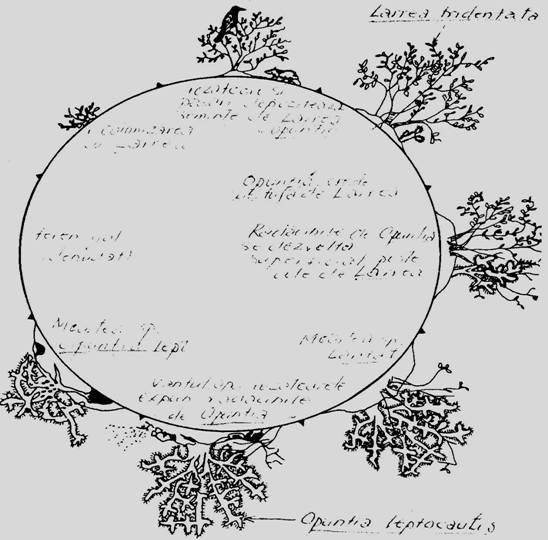
Fig. 6-58. Model sugestiv privind inlocuirea ciclica dintre speciile de desert Larrea tridentata si Opuntia leptocaulis din desertul Chihuahuan (dupa Yeaton, 1978).
5. Fluctuatii
Reprezinta o categorie de modificari si transformari la nivelul biocenozelor, o alterare a structurii si compozitiei biocenozei, ca urmare a unor dezechilibre puternice la nivelul factorilor de mediu (furtuni, uragane, seceta, inundatii). In urma fiecarui eveniment, nu toate speciile se comporta la fel. In functie de durata si intensitatea factorului perturbator este si amploarea modificarilor din biocenoze. Daca actiunea este mai slaba si nu are loc decat o distrugere partiala, biocenoza si ecosistemul se refac si chiar daca unele specii sunt inlocuite cu altele, structura comunitatii se reintoarce la "normal". Pentru intervalul redus an care sunt prezentate imaginile, nu este vorba de o succesiune reala.
Daca fluctuatiile sunt puternice si de durata ecosistemul vechi "moare" (si implicit biocenoza) si in timp locul sau este preluat de o biocenoza noua, de un nou ecosistem. Are loc o succesiune.
6. Succesiunea ecologica
Orice sistem din natura are o durata de viata limitata si este marcat de o evolutie ireversibila. Biocenoza si ecosistemul se incadreaza in aceasta categorie. Succesiunea ecologica este asadar o schimbare ireversibila in structura biocenozelor, a ecosistemelor, care determina inlocuirea unui ecosistem cu altul. Si succesiunile ecologice au la baza schimbarea factorilor fizici, iar ca urmare a acestora are loc o puternica interactiune intre componentele biocenozei, intre parametrii structurii functionale.
Cu alte cuvinte, succesiunea ecologica este un complex de interactiuni de distrugere a structurii si functiilor existente intr-o biocenoza sau ecosistem si formarea de noi structuri si functii.
Succesiunea ecologica este o inlocuire sistematica si ordonata a unei comunitati de catre o alta pina ce comunitatea atinge o relativa stabilitate in acel areal
In cadrul succesiunii exista o serie logica de etape care se deruleaza una dupa alta (de la stadiul de vegetatie ierboasa la tufaris si apoi padure) iar in final se termina cu o comunitate stabila. Ansamblul de etape se cheama serie, iar orice schimbare ce poate avea loc este denumita stadiu al seriei. Fiecare stadiu al seriei este in sine o comunitate, alta fat[ de cea anterioara si care poate ramane asa, o perioada scurta sau mult mai lunga de timp.
Este interesant de precizat ca initial a existat un punct de vedere geobotanic dupa care, succesiunea ecologica este doar o inlocuire a covorului vegetal de pe un anume teren, disparitia vechilor asociatii, geneza altuia nou si evolutia noilor asociatii vegetale. Ulterior conceptul a fost interpretat ecologic (Tischler, 1955).
Din punct de vedere ecologic, succesiunea ecologica este o degradare, inlocuire, geneza si o evolutie a asociatiilor vegetale, animale si de microorganisme, o evolutie a biotopului, si a unor structuri trofodinamice si spatiale, variatii ale mediului fizic, climatice si geomorfologice.
Succesiunea ecologica decurge din structura biocenozei, a ecosistemului, din interactiunile dintre specii, din metabolism si nivelul allelopatic. Toate acestea creaza o tensiune ecologica, contradictii interne intre componente, procese evolutive si remaniere structurala (Stugren, 1975). Chiar daca decurge din insasi structura ecosistemului, succesiunea este declansata de factori esterni (Bormann si Likens, 1979).
Trecerea de la o faza la alta (de ex. de la pajiste la teren acoperit cu arbusti) este insotita de pierderi de energie si productie crescuta de entropie (Margalef, 1962).
Chiar si atunci cand nu au loc catastrofe ale mediului fizic, are loc o inlocuire de lunga durata a biocenozelor si ecosistemelor deoarece chiar dinamica relatiilor interspecifice duce la transformari si modificari structurale permanente. Inlocuirea speciilor care edifica o biocenoza existenta, veche, cu altele noi, concomitent cu transformarea in timp a conditiilor noi de existenta se numeste succesiune (Clements, 1928).
6.1. Succesiunea primara (ecogeneza). Teoria clasica si teoria stohastica a succesiunii
Atunci cand noua biocenoza si noul ecosistem se formeaza pe un teren care initial era abiotic (lipsit de viata), este vorba de o succesiune primara. Este de exemplu cazul unei insule dupa o eruptie vulcanica. Se da ca exemplu insula Krakatoa (Indonezia), distrusa complet de o eruptie vulcanica in 1883 si unde s-a dezvoltat a noua padure tropicala dupa 30 de ani. Uneori procesul este si mai indelungat.
Noua structura a biocenozei, a ecosistemului, nu se bazeaza pe o biomasa dintr-o structura anterioara ci se alimenteaza din exterior, prin specii care initial ajung accidental in noul mediu din formatiuni biotice invecinate. Noile specii care colonizeaza si ordinea ierarhica depinde de natura geomorfologica a biotopului si de noile conditii de mediu. Se schimba si natura chimica a interrelatiilor dintre membri biocenozei.
Ecogeneza (geneza si evolutia biocenozelor si ecosistemelor) nu este o simpla succesiune de specii intr-un biotop ci un proces evolutiv prin care, masa biotica si biotopul creaza structuri ecologice noi, prin interactiune (noi sisteme de interrelatii intre viata si mediu). Deci ecogeneza include: succesiunile vegetatiei, faunei, geomorfogeneza si formarea solului.
Ecogeneza se suprapune cu conceptul de succesiune primara, adica cu istoria naturala a dezvoltarii unei biocenoze de la pionierat (colonizarea primara a spatiului abiotic) si pana la stadiul de stabilizare (climax) (Stugren, 1994).
Teoria clasica a succesiunii ecologice primare. Elaborta initial (Clements, 1928) si ulterior completata (Rickleffs, 1976), teoria clasica a succesiunii ecologice primare este o teorie determinista care o dominat gandirea ecologica a vremii. Constituirea noii biocenoze este rezultatul unei succesiuni legice dependenta de mecanismele interne ale dinamicii biocenotice Astfel, fiecare compozitie de vegetatie si fiecare stadiu actioneaza permanent asupra solului si constituie premisele infiintarii unui nou fond vegetal. Cu alte cuvinte sunt asigurate permanent premisele ca sa existe o succesiune iar compozitia stadiilor sa se succeada dupa ordinea filogenetica a clasificarii plantelor in sistemele naturale de clasificare. Plantele superioare nu vor putea actiona ca pionieri pe un sol fara vegetatie incipienta sau existenta, iar acest lucru poate fi realizat doar de plantele inferioare (ex. de ordine fireasca: alge albastre-----briofite-----pteridofite-----gimnosperme-----angiosperme). Aceasta teorie determinista este sustinuta de istoria naturala a reimpaduririi insulei Krakatoa (1883). Admitandu-se ca intreaga viata de pe insula a fost distrusa, reimpadurirea naturala a insului s-a realizat in circa 3 decenii, dupa ordinea prezentata mai sus. Si in alte zone s-a observat ca pionierii succesiunii ecologice sunt algele albastre (Marathe si Caudhari, 1975).
Dar faptul ca logic, dupa aceasta ar aparea muschii si pteridofitele si nu plante superioare, a fost o supozitie si nu o legitate dovedita.
Cercetari ulterioare au dovedit si alte
aspecte. Astfel, in cazul unor vulcani (
In ceea ce priveste lumea animala datele sunt si mai ciudate. Chiar pe insula Krakatao se pare ca unele animale au aparut pe insula chiar inainte plantelor sau cel putin odata cu ele. Chiar aici se pare ca primul animal care a imigrat din insulele vecine a fost un sarpe piton (Python reticulatus). La un an dupa eruptie au aparut pe malul marii colembole, diptere si coleoptere, iar acestea au atras pasarile.
Initial in cadrul acestei teorii s-a sustinut ca pot exista fenomene intimplatoare in succesiunea ecologica dar acestea sunt valabile doar pentru faza de pionierat, cind spori, polen, seminte, microfauna sunt aduse neselectiv de vant (sau ape). Dupa aceasta insa, selectia naturala a factorilor locali de mediu va induce un proces evolutiv ordonat, fiind mentinute doar acele organisme care sunt adaptabile la noile conditii de mediu. Din momentul in care intervine eceza (colonizarea) biocenoza nu se va mai dezvolta la intamplare ci ca rezultat al interactiunilor dintre specii, determinate de factorii ecologici.
Conform celor de mai sus se poate constata ca teoria are la baza pe langa date observabile, mult mai multe deductii logice, ipoteze si predictii.
Astfel, nu s-a tinut cont ca orice biocenoza in evolutia sa se bazeaza pe strategii de adaptare a speciilor la noile conditii de mediu si doar daca fluctuatiile acestor conditii sunt extreme pot fi inlaturate specii care nu reusesc sa se adapteze (Hulst, 1978). Daca un anumit stadiu in dezvoltarea unei biocenoze este inlocuit cu un altul, fenomenul nu este rezultatul unei legi naturale predeterminante care trebuie sa se deruleze pas-cu-pas, ci este raspunsul pe care il dau organismele vii in urma interactiunilor lor cu factorii ecologici si nu in ultimul rand, bazat pe realizarea unor noi legaturi determinate de mediatorii chimici (Rice, 1974).
Teoria stohastica (moderna) a succesiunii ecologice primare. Are la baza datele observabile an studiul reimpaduririi unor insule unde viata a fost nimicita de eruptii vulcanice catastrofale, insule aparute din mare (ex. insula Surtsey, 1963), alte biotopuri nude din interiorul continentului determinate de diversi factori distrugatori. In toate aceste situatii nu a fost observata vreo succesiune legica a stadiilor de vegetatie ci totul s-a bazat pe o imigrare necontenita si intamplatoare. Din acest punct de vedere, cercetarile moderne au dovedit ca ecogeneza (succesiunea primara) este un fenomen stohastic (aleator, intamplator) care urmeaza nu o lege naturala predeterminata ci o lege probabilistica.
Modelul ecogenezei pe insula Surtsey este extrem de sugestiv. Pe spatiul abiotic constand din cenusa vulcanica (tefra) a inceput colohizarea microbiana in 3 faze: preecogenetica; faza de biotopuri de pionierat si faza de ecogeneza difuza (aut. cit.in Stugren, 1994). Planctonul a ajuns pe insula chiar cand aceasta era improprie vietii (gaze, substante toxice, lipsa apei, temperaturi foarte mari). In primul an s-au instalat bacteriile si algele formind "oaze de ecogeneza". Au aparut apoi si seminte de plante vasculare. Faza de pionierat s-a repetat de mai multe ori deoarece valurile mari si furtunile au indus alte fenomene de distrugere. Dupa 5 ani de la dezastru s-a inregistrat prima planta cu flori (Cakile maritima). Briofitele au aparut mai tarziu si au acoperit solul gol. Au aparut acum si artropodele [gr. arthros = articulat; poos, podos = picior]. La 7 ani de la dezastru pe insula erau inventariate: o flora spontana cu 7 specii si o fauna asociata de 5700 indivizi de artropode, apartinand la 158 de specii (majoritatea diptere). Insectele au ajuns pe insula tot gradat, in mai multe valuri, prin zbor propriu sau aduse de curenti. Imigrarea a fost mai rapida decat colonizarea.
Un alt aspect ce trebuie semnalat. Cu cat ecogeneza are loc la latitudini mai nordice cu atat durata este mai mare. Este dat ex. insulei Krakatoa unde dupa 50 de ani de la catastrofa deja exista padure pe insula, iar la 70 de ani padurea era compacta, pana in varful craterului. In zona subarctica insa (Kamciatka) padurea s-a dezvoltat doar dupa 250 ani iar pe zona pe unde s-a scurs lava, padurea s-a constituit dupa 800-1200 de ani.
Un caz interesant de succesiune primara este la nivelul zonei subalpine. In stadiile timpurii, pe stincile goale (sau denudate) vor imigra speciile pionier (alge, licheni, muschi), proces continuat in timp de colonizarea altor specii. In faza urmatoare, prin concurenta interspecifica se afirma speciile lemnoase (ex. Picea excelsa). Pana va ajunge la stadiul de stabilitate care va fi o comunitate ierboasa mixta se trece si prin stadiul de tufaris inalt (Fig. 6-59).

1 2 3 4 5 6 7
Fig. 6-59.
Model de succesiune primara in zona subalpina (caz din
6.2. Succesiunea secundara si tipurile de succesiuni secundare
Atunci cand noua biocenoza se constituie dintr-una existenta, din ramasitele unui ecosistem partial sau total distrus (ex. microorganisme, nevertebrate care se ascund in sol, radacinile profunde ale plantelor, organe vegetative din sol - bulbi, rizomi, tuberculi etc), se vorbeste de o succesiune secundara sau simplu succesiune. Acestea (spre deosebire de succesiunile primare) apar peste tot in biosfera. In acest caz, pentru declansarea lor nu este nevoie de un teren complet nud ci doar de interventia unor actiuni perturbatoare (furtuni, incendii, inundatii, modificari in regimul trofic sau chimic al habitatelor, interventia factorului antropic). Dupa cum am aratat mai sus (vezi fluctuatiile) aceste actiuni pot fi urmate doar de modific[ri cantitative a elementelor floristice si faunistice componente, a variatiei (fluctuatiei) numarului de specii sau indivizi din biocenoza respectiva. Daca actiunea este mult mai accentuata si de durata, se ajunge la succesiune ecologica datorita faptului ca ordinea veche din biocenoza se modifica si se inlocuieste cu o alta, in acelasi habitat.
Dupa natura, amploare si intensitatea actiunii ce o declanseaza se cunosc mai multe tipuri de succesiuni: autotrofe, heterotrofe, abiogene, biogene, tehnogene, microsuccesiuni, succesiuni experimentale.
a. Succesiuni autotrofe. Este cel mai comun model de succesiune din natura. Se numeste astfel deoarece incepeintr-un mediu predominant anorganic si este caracterizata de dominanta timpurie cit si ulterioara a plantelor autotrofe. Sunt de fapt cele mai comune. Poate fi cazul succesiunii unei dune de nisip spre vegetatie lemnoasa (Fig. 6-60) sau a unei zone intens pasunate spre padure (Fig. 6-61).

Fig. 6-60. Succesiunea dunelor de nisip. De la o duna localizata la marginea unui lac si acoperita cu o specie ierboasa (A), printr-un amplu proces de inmultire si propagare a acestora (B) dunele se "fixeaza" la distanta mai mare de lac (C) iar in final dunele batrane sunt acoperite de vegetatie lemnoasa (dupa Brewer, 1988).

Fig. 6-61. O suucesiune tipica pe o suprafata initial afectata prin pasunat intensiv (a); aceeasi zona dupa 21 ani (b); dupa ce portiunea din zona a a fost ingradita, vegetatia s-a modificat profund dupa 20 de ani (d) (dupa Smith, 1974).
b. Succesiuni heterotrofe. Acest model de succesiune este caracterizat de dominanta timpurie a speciilor heterotrofe (animalele) si se deruleaza acolo unde biotopul este initial de tip organic.
c. Succesiuni abiogene. Pornind de la un substrat biotic initial existent, se formeaza biocenoze si ecosisteme noi datorita actiunii factorilor fizici (abiotici), fie ca sunt actiuni moderate (modificari climatice) fie catastrofale (cutremure, inundatii, vulcani, incendii, uragane, avalanse).
Transformarile climatice, spre deosebire de celelalte, nu determina intotdeauna schimbarea biocenozelor ci doar inlocuirea unor specii, fara ca astfel sa se modifice substantial planul initial de structura (ex. deoarece climatul Europei centrale si nordice a devenit tot mai oeanic aceasta a dus la imigrarea de specii de pasari iubitoare de umiditate, in tarile din nord, paralel cu disparitia altor specii care existau acolo) (Curry-Lindahl, 1961). De asemenea, modificarile climatice determina succesiuni lente, de lunga durata (ex. este cazul amestecului straturilor de apa din zona litoralului englez in perioada anilor 1950; ca urmare apa de litoral a devenit oceanica si la acest nivel a ajuns masiv si zooplanctonul pelagic si implicit pestele plantofag Clupea harengus).
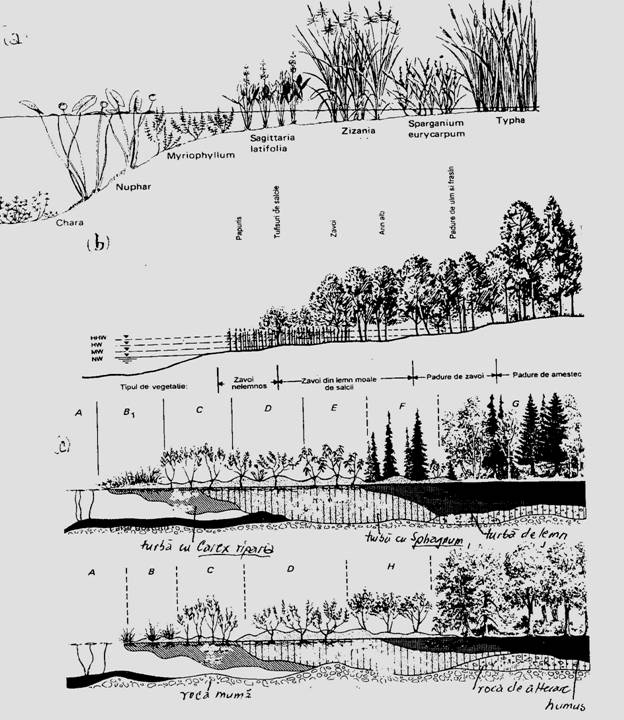
Fig. 6-62. Succesiunea biogena avatica. O secventa din succesiunea lac-----mlastina eutrofa, in faza initiala, cu instalarea macrofitelor pe marginea lacului (lipsesc fazele initiale: pionieratul algelor = planctonul, microorganismele si micronevertebratele); algele de tip Chara sp. formeaza primul tip de sediment pe care se vor instala speciile urmatoare ale succesiunii (a); succesiunea vegetatiei la malul unui rau (b); o evolutie lunga marcheaza succesiunea mlastinii eutrofe spre sfagnete si apoi spre padure, compozitia acesteia din urma fiind diferita in functie de fazele anterioare ale succesiunii (c) (dupa Dansereau si Sagadas-Vianna, 1952; din Smith, 1974; Godeanusi Paraschiv, 2005).
Actiunea catastrofala a factorilor de mediu este mult mai profunda. Eruptiile vulcanice prin emanatii de cenusa, lava si gaze pot modifica semnificativ mediul si implicit structura biocenozelor (ex. eruptia vulcanului St. helen - SUA, prin cenusa care a ajuns in apa a determinat eutrofizarea lacurilor de munte, s-a accelerat metanogeneza si activitatea bacteriilor sulfooxidante iar metabolismul lacului a devenit asemanator cu activitatea metabolica a estuarelor).
Incendiile ca si calamitati naturale, apar
frecvent in anumite zone (savanele est-africane, padurile de pin din
d. Succesiuni biogene. Daca exista conditii abiotice relativ constante atunci succesiunea ecologica este determinata de concurenta interspecifica (ex. este vorba de o succesiune acvatica cu cel putin doua directii de succesiune: succesiunea de la lac la mlastina eutrofa, urmata de trecerea de la o mlastina eutrofa cu macrofite pe margine, la un tinov oligotrof cu Sphagnum (Pop, 1960), concomitent cu inlocuirea faunei vechi si aparitia de noi combinatii de specii) (Fig. 6-62, a, b).
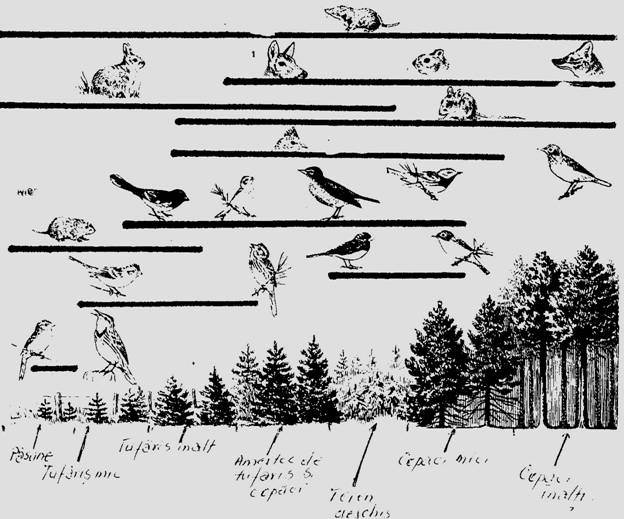
In cazul succesiunii biogene exista si o succesiune de combinatii de specii care se succed dpendent de natura vegetatiei si solului (Fig. 6-63). In biotopurile primare spectrul formelor biotice de animale este bogat dar se va restrange progresiv pe masura ce succesiunea avanseaza (ex. recifele coraliere sunt locuite si atacate de o serie de specii minatoare - spongieri, bivalve, polichete, sipunculide, ciripede, etc - iar resturile se vor depune; pe acestea se colonizeaza alge apoi fanerogame marine - Thallasia sp. - iar apoi coralierii - Acropora sp., Millepora sp.) (Margalef, 1962).
(a)
(b)
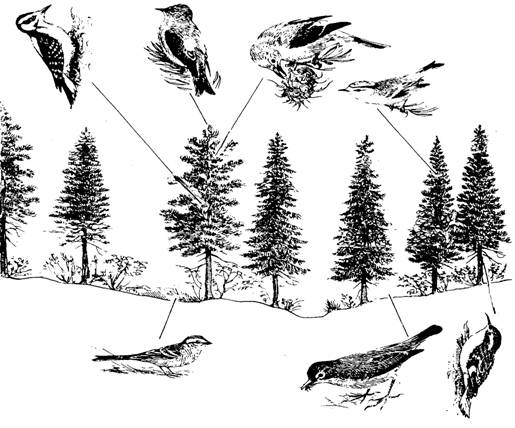
Fig. 6-63. O succesiune biogena la nivelul unei plantatii de conifere. Liniile orizontale marcheaza teritoriul ocupat de respectivele specii de vietuitoare imigrante (a). Unele specii vor dispare, fiind inlocuite cu altele, iar altele vor ocupa suprafete extinse iar in final, in faza de climax vor avea un spatiu vital (nise) bine definit (b) (dupa Smith, 1974).
Realizarea unei succesiuni biogene este dependenta de capacitatea plantelor (si in special a semintelor) si animalelor imigrante de a intra in biotopi deja ocupati de alte vietuitoare, de a determina izgonirea lor de aici si de a ocupa biotopul. Nu exista nici o legitate, deoarece conditiile de mediu sunt fluctuante si in acest caz colonizarea este aleatoare (stohastica
Concurenta interspecifica a influentat si extinderea culturii de Saccharum officinale (trestia de zahar (in insulele Ryukyu a fost introdusa prin jurul anului 100 si ocupa in prezent aprozimativ 5000 ha; in Cuba a fost introdusa mult mai tarziu - 1540 - dar astazi ocupa aproximativ 950.000 ha; concluzia: in insula Ryukyu erau doar 21 specii de insecte daunatoare iar in Cuba 81 de specii).
e. Succesiuni tehnogene. Sunt determinate de actiunile economice dirijate ale omului (prelucrarea solului - agricultura, industrializarea, incendierea intentionata, modificarea voita a habitatelor naturale - baraje, lacuri, sosele, cai ferate, viaducte, constructii industriale, cariere, deseuri industriale, halde de steril etc). Influenta de durata a omului asupra biocenozelor naturale si asupra ecosistemelor naturale, modifica mai mult sau mai putin, voluntar sau involuntar, conditiile de existenta ale animalelor si plantelor si induce aparitia succesiunilor. Se considera de catre unii ca si omul primitiv, prin agricultura promitiva practicata si prin vanat ar fi determinat succesiuni ecologice. Mai nou se pare improbabil acest lucru deoarece omul primitiv sau oamni care actualmente au un mod de viata primitiv (triburile) si care traiesc in mijlocul naturii, manifesta o mult mai mare atentie fata de aceasta si o "intelege" mult mai bine (ceea ce pare un paradox daca ne gandim la omul modern si cultivat). Este cazul culturii traditionale intinerante a solului (Fig. 6-64).
Influenta factorului antropic asupra succesiunii ecologice are o serie de exemple edificatoare. Extinderea suprafetelor pentru agricultura si fanate a determinat extinderea arealului unor vertebrate ca Alces alces (elanul) si Capreolus capreolus (capriorul), dar au indus dezavantaje totale pentru o serie de alte specii cum ar fi Galerida cristata (ciocarlanul), Ciconia ciconia (barza alba), Bombina bombina (buhaiul de balta).
Sunt interesante succesiunile care au loc pe terenurile agricole care sunt lasate in paragina. Corelat cu vegetatia generala a regiunii, in timp se va evolua spre padure sau spre stepa.
Daca se defriseaza un teren forestier prin succesiune se va ajunge din nou la padure. Imigrantii, vegetali si animali, vor proveni din zonele invecinate, nedefrisate.
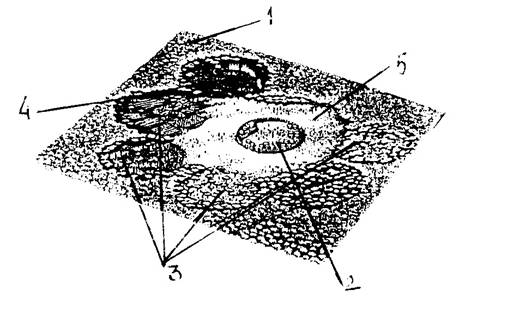
Fig. 6-64. Ilustrarea modelului cultivarii intinerante a pamintului de catre anumite triburi. 1 - padurea compacta; 2 - pozitia satului; 3 - suprafete circulare care au fost initial defrisate prin ardere controlata; in aceasta suprafata ce corespundea cam 1 ha/50 locuitori s-a practicat agricultura de existenta; dupa ce solul s-a epuizat, s-a trecut la defrisarea unei suprafete alaturate iar cea anterioara a fost plantata cu puieti pentru impadurire; dupa un circuit complet, in prima suprafata padurea este refacuta; 4 - ultima suprafata defrisata prin ardere (din Larousse, 1993).
Actiunile voite ale omului sunt frecvent deosebit de distructive si interventia se lasa cu succesiuni ecologice, uneori atipice. Am mai amintit ca organismele din varful piramidelor trofice au o pozitie de "sef conducator". Este o pozitie avantajata in mecanimul de feed back deoarece actiunea lor asupra nivelului trofic imediat inferior (C2) determina reactii in lant iar efectul se repercuteaza in toate verigile lantului trofic si chiar in nodurile retelelor trofice. Se induce astfel o dezorganizare a ordinii existenete ce poate finaliza cu denudarea biotopului respectiv. Din acest moment incepe succesiunea. Daca in virful piramidei trofice este omul, el se va comporta asupra verigii imediat inferioare ca si un carnivor de varf asupra consumatorilor secundari. Efectuarea unui baraj determina modificari profunde ale biotopului, florei si faunei (Fig. 6-65).
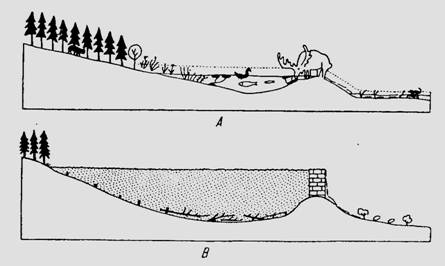
Fig. 6-65. Efectul negativ al indiguirii unui curs de apa asupra diversitatii biologice (dupa Dansereau, 1957).
Extrem de sugestiva este redata influenta factorului antropic asupra mediului, de-a lungul existentei sale (Fig. 6-66). S-a ajuns de la padurea virgina, sustinatoare a vietii, la mediul urban, sofisticat, dirijat si stresant.

Fig. 6-66. Reprezentare schematica sugestiva a actiunii factorului antropic asupra unui ecosistem natural (padurea), la nivelul diferitelor perioade de timp (dupa Dansereau, 1957). A. padure virgina; B - etapa de culegator; C - etapa - vanatoare; D - pastoritul; E - agricultura; F - industria; G - urbanizarea.
Pe de alta parte, actiunile omului pot sa fie facute si in avantajul naturii. Astfel este edificatoare interventia sa in creerea haturilor dintre parcelele cultivate, impadurirea malurilor unui lac, "renaturalizarea" malurilor unui parau etc (Fig. 6-67, a, b), actiuni cu rol deosebit de important in cresterea biodiversitatii. Aceste actiuni controlate si supravegheate stiintific se constituie in etape importante in procesul de restaurare ecologica.
(a)

(b)
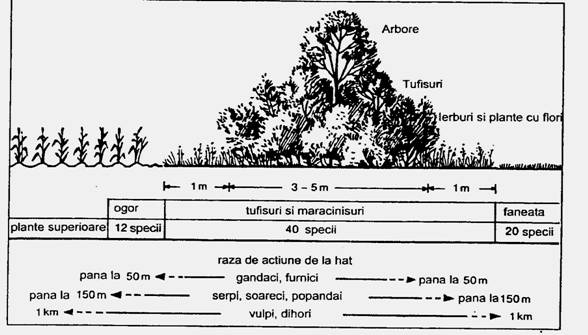
Fig. 6-67. Succesiuni tehnogene. Interventia omului in dirijarea succesiunilor: constituirea haturilor ca o retea care uneste spatiile naturale (a); la nivelul unui hat exista o vegetatie caracteristica iar fauna instalata aici poate actiona la diferite distante (b) (dupa Dingethal si colab., 1995; Godeanu si Paraschiv, 2005).
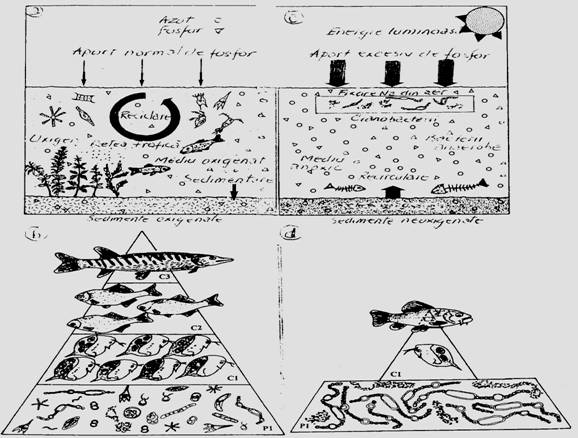
Fig. 6-68. Functionarea normala a unui lac (a) si piramida trofica reprezentativa a acestei stari (b). In conditiile unui lac eutrofizat (c), priramida trofica este anormala (d) (dupa Devaux, 1976; Barroin, 1991; din Faurie si colab., 1998).
Una din succesiunile tehnogene interesante este fenomenul de eutrofizare, cel mai frecvent, rezultat al poluarii. In conditii normale de functionare, un lac de exemplu acumuleaza o cantitate normala de azot si fosfor. El functioneaza normal si piramida trofica este bine reprezentata (Fig. 5-91, a, b). Prin eutrofizare, ca urmare a acumularii unei cantitati mari de azot si fosfor, echilibrul se rupe, organismele mor si piramida trofica devine anormala (Fig. 6-68, c, d).
Prin biomanipularea biotopului si a organismelor componente ale piramidei trofice sau a retelelor trofice, omul poate interveni in diminuarea sau inlaturarea eutrofizarii.
f. Microsuccesiuni (microserii) (Mohr, 1943; Shelford, 1951). Tot ceea ce am amintit anterior sunt practic succesiuni derulate pe suprafete mari (macrosuccesiuni). Exista insa si succesiuni ecologice ce se desfasoara in microbiotopuri si unde au loc aceleasi fenomene in care o anumita cenoza existenta, prin modificarea conditiilor de viata, va fi inlocuita cu o alta noua. Acestea sunt microsuccesiuni ecologice. Asemenea fenomene au loc in luminisuri de padure sau in conditii nou create printr-o interventie initiala a omului.
g. Succesiuni experimentale. Este dificil de a organiza un experiment controlat in conditii de laborator, a unui fenomen de succesiune ecologica. Experimentele au evidentiat fenomenul doar la nivelul microorganismelor si al unor specii de nevertebrate. Pentru specii de nevertebrate s-au creat "insule artificiale" (panouri sterile din lemn) care s-au dispus in ecosisteme acvatice marine la diferite adancimi. Dupa cca. Un an toate "insulele" au ajuns, la un numar egal de specii, indiferent de pozitia geografica (la cele situate spre latitudini mai sudice colonizarea s-a realizat in timp mai scurt), ceea ce dovedeste ca este hotaratoare concurenta intraspecifica si nu alt factor (Schoener si colab., 1978). Pe linga microorganisme s-au instalat consumatori ai lemnului, pe urma consumatori ai lemnului putrezit iar detritusul organic a fost atacat de speciile de saprofage. Rol important au avut nevertebratele marine imigrante. Concurenta a influentat care specie este stabila. Furtunile au distrus o parte din specii, indiferent de nivel biotic iar supravietuitorii au scapat datorita intamplarii si nu ca urmare a "luptei pentru existenta".
Un alt exoeriment derulat intr-o mlastina de mangrove in SUA, unde o anumita suprafata s-a sterilizat prin gazare si unde s-a instalat apoi tot o "insula" naturala a aratat ca succesiunea a decurs ca un proces stohastic.
Aceste modele experimentale nu pot insa reda amploarea si realitatea succesiunii ecologice reale. Prin modelare matematica si reprezentare teoretica se poate insa sugera amplitudinea evenimentelor si activitatilor in derularea fazelor succesiunii (Fig. 6-69).
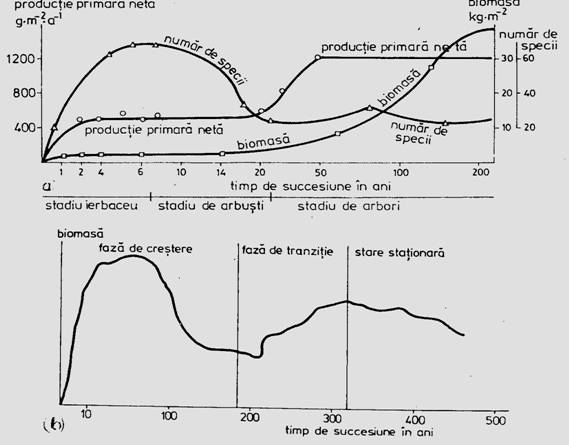
Fig. 6-69. Ilustrarea teoretica a succesiunii primare a unui paduri (a) si evolutia numarului de specii, productiei primare si biomasei in ecogeneza padurii (b) (dupa Bormann si Likens, 1979; Whittaker, 1975; din Stugren, 1994).
6.3. Stadiile (fazele) si trasaturile succesiunii ecologice
Clements (1928) si cercetarile ulterioare au evidentiat existenta a 6 stadii succesive: 1 - denudarea biotopului; 2 - imigrarea; 3 - eceza (colonizarea) [gr. oikizo = eu colonizez]; 4 - concurenta interspecifica; 5 - reactia biocenotica; 6 - stabilizarea.
1. Deteriorarea si distrugerea biocenozelor originale si a ecosistemului (denudarea biotopului). Biocenoza sau ecosistemul, ca structuri ecologice, sunt elemente fragile ce pot fi usor perturbate de factorii de mediu, biotici sau abiotici.
Factori abiotici ce determina distrugerea unei biocenoze sau a unui ecosistem (Schafer, 1962): evenimente geomorfologice ce schimba configuratia habitatului (alunecari de teren, vulcanism, cutremure, incendii, inundatii); sedimentarea pe fundul ecosistemelor acvatice a unor substante; acumularea de substante toxice etc
Factori biotici de distrugere: boli, pradatori care au imigrat in numar foarte mare; invazia de specii straine cu potential bitic mult mai mare decat al speciilor indigene; autointoxicarea mediului; acumularea de organisme moarte care nu sunt rapid descompuse (tanatocenoze) etc.
2. Formarea noii biocenoze si a noului ecosistem. Denudarea biotopului este urmata de fenomenul imigrarii (pionierat) care a fost abordat mai sus. Se refera la organismele capabile sa transforme terenul abiotic intr-un habitat unde viata este posibila. In faza de pionierat, factorul decisiv este deci hazardul.
In faza de colonizare (eceza) intervine selectia naturala a factorilor fizici ai mediului. In faza concurentei interspecifice (competitia) intervin factorii biotici, interrelatiile directe dintre specii.
Reactia biocenotica consta in existenta "unui plan comun" al speciilor ca reactie uniforma a lor fata de mediul de viata si implicit o transformare a mediului sub actiunea masei biotice.
Atunci cind biocenoza este suficient de matura si stabila iar ecosistemul are deja "personalitate" si autonomie fata de mediu se atinge de fapt faza finala - stabilizarea (climax).
Succesiunea ecologica are urmatoarele trasaturi:
☻ - in cursul succesiunii, biomasa, productia si producivitatea cresc;
☻ - in cursul succesiunii creste snumarul de lanturi trofice si se constituie noi retele trofice;
☻ - in decursul succesiunii se trece progresiv de la structuri simple la structuri diversificate;
☻ - succesiunea este un proces discontinuu, de inlocuire a speciilor in acelasi biotop iar noile specii care vor determina noua structura provin dominant din exteriorul vechii structuri, prin imigrare;
☻ - pe plan energetic, succesiunea incepe cu mari cheltuieli de energie si tinde spre o reducere a risipei de energie (reducerea entropiei);
☻ - succesiunea este o secventa a unei stari de echilibru dinamic (fluent).
Succesiunea ecologica cuprinde si o serie de stadii botanice in diferite conditii climatice (Clements, 1928):
- xeroserie - daca dezvoltarea vegetatiei se desfasoara de la un spatiu abiotic arid la unul mezofil (ex. da la camp arid la padure) (Fig. 6-70, a);
- hidroserie - daca dezvoltarea vegetatie are loc de la un mediu umed spre un biotop arid (ex. de la o plaja inundata spre un loc arid (Fig. 6-70, b).
6.4. Climax
Climax (Clements, 1928) semnifica un stadiu final, culminant, in evolutia unui proces. El nu inseamna "moartea" ecosistemului si sfarsitul evolutiei ci doar o atenuare a vitezei de evolutie (macrosuccesiunea este inlocuita de microsuccesiuni, in diferite puncte ale ecosistemului). In cazul succesiunilor ecologice, climaxul este starea finala spre care tinde biocenoza, ecosistemul, o stare stationara de durata care se mentine stabila timp indelungat (ex. codrii seculari din valea Cernei, de la Slatioara din Masivul Rarau, padurea de stejar de la Letea si Caraorman etc). Astfel, sensul evolutiv al unei biocenoze s-au ecosistem este spre o stare de echilibru cand se realizeaza homeostazia [gr. homoios = asemanator; stasis = stare]. Aceasta este valabil atat timp cat nu intervine un factor perturbator violent.
Stadiul de climax [gr. klimax = scara] implica un timp indelungat pentru instalarea sa (chiar si in regiunea tropicala unde o padure se reface in 40-50 de ani, climaxul se instaleaza mult mai tirziu; dupa parerea specialistilor un ecosistem se stabilizeaza definitiv structural dupa intre 500 si 4000 de ani !).
Trebuie retinut ca stadiul de climax este o stare stationara in evolutia unui ecosistem si nu este vorba despre maturitatea unui organism. In lucrari de inceput terminologia clementsiana a climaxului uza si de alte notiuni asupra carora nu vom insista (proclimax, subclimax, serclimax, disclimax, preclimax, postclimax) (Brewer, 1988).
Exista 3 teorii importante care explica acest concept:
☻ - Teoria monoclimaxului (climaxul climatic) (Clements, 1928). Acest stadiu de climax este atins atunci compozitia si structura vegetatiei sunt armonizate fata de un singur factor al mediului - clima. Conceptul este privit din punct de vedere al vegetatiei - geobotanic.
☻ - Teoria policlimaxului (Du Rietz, 1930). Structura vegetatiei se stabilizeaza corelat cu clima si solul. Astfel pe langa climaxul climatic exista si un climax edafic.
☻ - Teoria climaxului ecologic (teoria configuratiei de climax; conceptul modelului climax) (Whittaker, 1953). Conform acestei teorii, climaxul este o stare stationara, caracterizata si de o oarecare instabilitate dar cu un grad de variatie ce nu pericliteaza structura sistemului respectiv. Pentru a se initia instalarea climaxului, cel mai important aspect este stabilirea unor relatii stationare intre speciile care alcatuiesc biocenoza, o coadaptare functionala a populatiilor (Stugren, 1975).
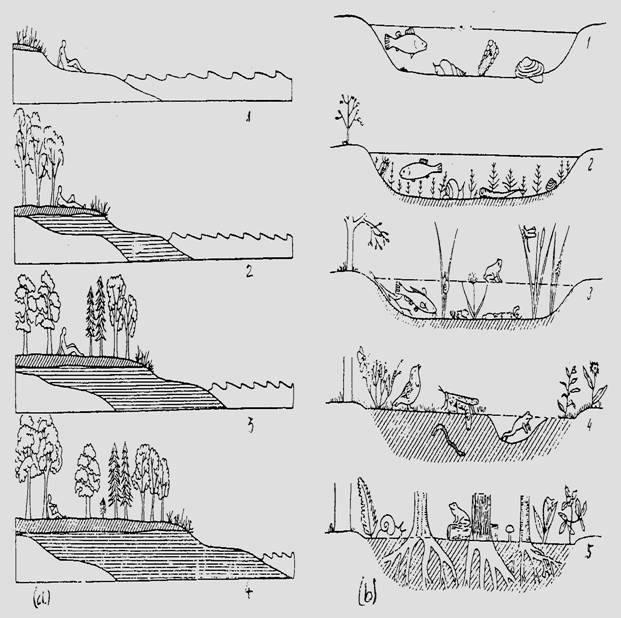
Fig. 6-70. Reprezentarea schematica a succesiunii ecologice in cazul unei xeroserii (a) si a unei hidroserii (b) (date din Allee si colab., 1949; Clarke, 1954; preluate din Stugren, 1965). Sunt figurate doar etapele mari, semnificative ale evolutiei in succesiune. a - nisip; b - nisip si praf adus de vant si valuri; c - humus nou format; 1 - pionierat; 2 - vegetatie submersa; 3 - vegetatie emergenta; 4 - dominarea suprafe'ei acoperita de campie si exiustenta unei balti reduse la minim; 5 - padure.
7. Succesiunea geologica
Succesiunile geologice sunt succesiuni ecologice (un lant de succesiuni primare si secundare) care s-au derulat de-a lungul a sute de mii sau milioane de ani. Succesiunile ecologice de azi sunt doar verigi ale succesiunii geologice, de durata mult mai scurta.
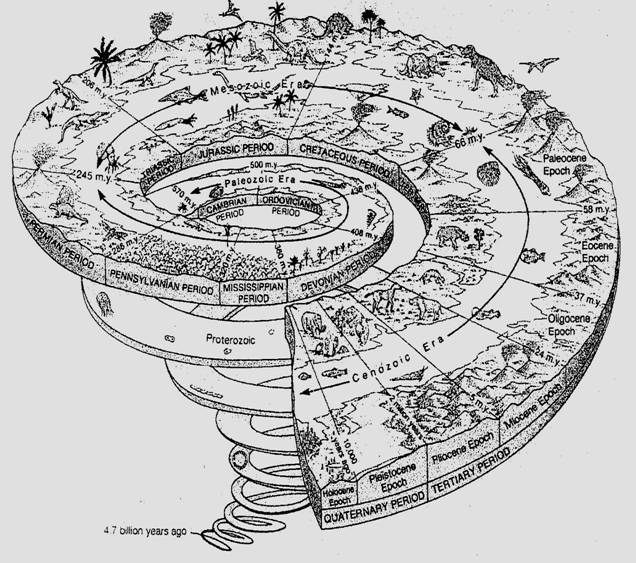
Fig. 6-71. Spirala timpului geologic. Sunt evidentiate "momente de rascruce" privind fenomenul succesiunii ilustrat prin cele mai importante grupe de vietuitoare.
Succesiunile primare (ecogeneza) de azi sunt rare, deoarece un fenomen de denudare completa a biotopului are loc rar. La scara geologica insa, Terra, la inceputurile sale a fost un imens biotop total dedudat si timp de milioane de ani scoarta terestra a fost o arena pe care s-au derulat nenumarate succesiuni primare. Faza de pionierat (dupa ce s-au constituit coacervatele din macromolecule organice si apoi organismele unicelulare primitive) au fost urmate de imigrare a organismelor in conformitate cu conditiile create in diferite biotopuri, apoi a fost cucerit mediul acvatic si in final cel terestru. Un proces de lunga durata, initiat prin precambrian si probabil terminat prin mezozoic sau chiar mai tarziu (Fig. 6-71). Pe un interval atat de extins se poate vorbi de o scara geocronologica a biosferei (Tabelul 15). In cadrul proceselor derulate in succesiunea primara, microorganismele ciupercile, plantele si animalele s-au adaptat la noile conditii aparute s-a dezvoltat speciati si implicit au aparut noi tipuri de biotopuri si de ecosisteme.
Tabelul 15
Scara geocronologica a geobiosferei si stratigrafia generala a Terrei.
|
EON (conotema) |
ERA (eratema) |
PERIOADA (sistema) |
EPOCA (sectia) |
DATAREA CU IZOTOPI (mil. ani) |
BIOSISTEM (fauna, biomuri si ecosisteme) |
|
Fanerozoic (faneros = vizibil) |
Kainozoic (tertiara) |
Cuaternar (antropogen) |
Holocen Pleistocen |
Plante si vegetatie |
|
|
NEOGEN |
Pliocen Miocen |
Evolutia veg. de tip actual |
|||
|
Paleogen |
Oligocen Eocen Paleocen | ||||
|
Mezozoic (secundara) |
Cretacic |
Tarziu Timpuriu |
Angiosperme Primele plante din grupa gimnospermelor Apar gimnospermele |
||
|
Jurasic |
Tarziu Mediu Timpuriu | ||||
|
Triasic |
Tarziu Timpuriu | ||||
|
Paleozoic (Primara) |
Permian |
Tarziu Mediu Timpuriu |
Ferigi si pteridofite |
||
|
Carbonifer |
Paduri tropicale; primele pl. cu seminte |
||||
|
Devonian | |||||
|
Silurian |
Domina psilofitinele si briofitele |
||||
|
Ordovician |
Alge verzi si rosii |
||||
|
Cambrian |
Domina cianofitele |
||||
|
Kriptozoic (precambrian) |
STROMATA | ||||
|
proterozoic |
|
|
Primele celule |
||
|
Inferior |
Mediu Inferior |
Molecule agregate Molecule Biofore |
|||
|
Arhaic |
>3500 |
Originea vietii |
|||
Succesiunea secundara a luat apoi locul celei primare, structura si compozitia ecosistemelor s-a remaniat iar pe diferitele nivele trofice ale niselor ecologice au aparut progresiv si treptat-treptat, noi specii. Astfel, odata ce a aparut o noua nisa ecologica oragnismele sale componente, bocenozele, nu au ramas neschimbate ci s-au succedat in timp conform principiului echivalentei geologice a componentelor biocenozei, nivelurilor trofice si niselor ecologice.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 10544
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2026 . All rights reserved