| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
IMUNITATEA PIELII
Pielea, tegumentul extern sau invelisul cutanat, ca organ de acoperire al corpului animal, exercita prin pozitia sa rolul de intermediar intre mediul extern si intern. Aceasta pozitie 'privilegiata' determina relatii functionale stranse cu organele protejate, carora le asigura mediul normal de activitate. La randul lor, organele protejate influenteaza integritatea morfologica si functionala a pielii. Dintotdeauna s-a insistat asupra rolului protector al pielii fata de numeroasele ingerinte din partea mediului extern prin factorii mecanici, fizici si biotici. Cercetarile ultimilor ani vin sa completeze rolul protector al pielii prin relevarea unei noi functii, aceea de organ imunitar (Fig.57)
Ca 'bariera a mediului extern', asemanator altor bariere, pielea este dotata cu o vascularizatie si o inervatie bogata, cat si cu sistem imunitar complex, capabil nu numai sa capteze informatiile antigenice, ci si sa le transmita organelor imunoformatoare si sa reactioneze promt in procesele de aparare locala si generala a organismului. Desi reactivitatea deosebita a stratului papilar al dermului (stratul cel mai reactiv din structura pielii) este cunoscuta de mai mult timp, abia din 1970, studiul markerilor si a functiunilor imunologice ale celulelor lui Langerhans a permis consacrarea pielii ca organ imunitar.
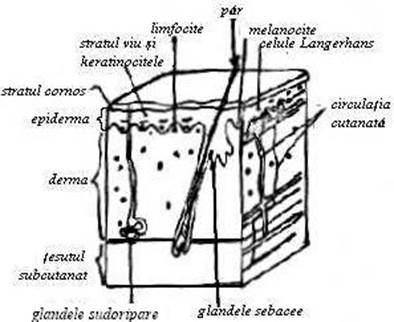
Fig. 57 Sectiune schematica prin piele
Cercetarile si observatiile clinice in legatura cu limfoamele pielii au sugerat ca aceasta ar avea o functie analoaga timusului.
Ipoteza legaturii stranse dintre timus si piele este confirmata de existenta, mai ales la animalele de laborator, a numeroaselor mutante care afecteaza concomitent pielea si timusul. Astfel, la soareci este cunsocuta mutanta nu. (nude mutation recissive), plasata pe cromozomul 11, care in stare homozigota determina absenta completa a parului si agenezie mai mult sau mai putin totala a timusului. Acesti soareci sunt incapabili sa matureze unele limfocite T. Ei pot fi partial normalizati prin transfuzie de limfocite T.
Separarea genelor responsabile de absenta parului de cele responsabile de agenezia timusului a fost imposibila, fapt care dovedeste ca cele doua seturi de gene sunt fie identice, fie strans legate pe acelasi cromozom.
Mutante care dovedesc absenta invelisului pilos si diferite leziuni congenitale ale timusului (pana la absenta totala a acestuia) au mai fost descrise la sobolani, cobai, hamsteri.
La soarece a mai fost descrisa mutanta bg (beige), gena mutanta recisiva plasata pe cromozomul 13, care antreneaza o profunda modificare a culorii pelajului si o diminuare importanta a rezistentei la infectia indivizilor homozigoti. Este echivalentul la soarece a maladiei Chediak-Higashi de la om. La sobolan este descrisa mutanta mK, indusa prin iradiere, caracterizata prin anomalii ale parului si o mare sensibilitate la infectii.
Studii electronomicroscopice au dovedit unele asemanari ultrastructurale intre timus si epiderm. Astfel, unele celule epiteliale timice sunt dotate cu structuri granulare specifice care par sa fie identice cu granulele din cheratinocitele epidermului. Cercetari cu anticorpi monoclonali fluorescenti au dovedit existenta unor cantitati de cheratina (proteina considerata proprie celulelor epidermice) in celulele epiteliale timice.
Un alt punct de legatura intre timus si piele este enzima dinucleotidiltransfareza, detectata in limfocitele ajunse intr-un anumit stadiu de maturatie in timus, dar care nu apare in limfocitele T mature. Daca limfocitele T sunt puse in cultura impreuna cu cheraticite, enzima isi face aparitia in cultura celulara.
Ulterior s-a dovedit interventia cheratinocitelor in maturarea partiala a limfocitelor T, prin intermediul unui hormon analog timopoietinei (hormonul timic cu rol principal in maturarea limfocitelor T). Cercetari cu anticorpi monoclonali fluorescenti au identificat pe suprafata celulelor epiteliale timice producatoare de timopoietina, trei receptori specifici TE4, A2Ba si q19. Acesti receptori au fost gasiti si pe suprafata membranei cheratinocitelor din stratul bazal al epidermului care produc hormonul asemanator timopoietinei. Un alt argument in favoarea rolului imunitar al cheratinocitelor ar fi faptul ca viteii cu paracheratoza congenitala au o susceptibilitate crescuta la infectii. La viteii respectivi se constata o hiperplazie a timusului.
Legatura functionala dintre piele si sistemul imun este sustinuta si de constatarea faptului ca o substanta exogena care patrunde in organism pe cale cutanata nu poate deveni imunogena, decat daca este prezentata limfonodurilor de catre limfocitele cantonate in tegument.
Substantele foarte imunogene, ca dinitrofluorbenzenul, aplicate pe tegumentul unui soarece care a fost expus in prealabil la radiatiile ultraviolete nu induc raspuns imunitar. Areactivitatea este remanenta si trasferabila la alt soarece prin transfuzie de limfocite T de la soarecele iradiat. Aceste date dovedesc ca pielea ar dispune de o clasa de celule care au proprietatea de a prezenta antigenul limfocitelor T-celule prezentatoare de antigen a caror functie poate fi anihilata prin expunere la radiatiile aultraviolete. Daca antigenul trece etapa acestor celule( scoase din functie prin iradiere cu ultraviolete) el induce o areactivitate remanenta, datorita stimularii limfocitelor T supresoare.
Investigatiile facute in scopul identificarii celulelor implicate in acest proces s-au oprit la celulele lui Langerhans. Sunt celule imunocompetente provenite din maduva osoasa si rezidente in epiderm, unde reprezinta 2-4% din populatia locala. Nu exista o coloratie de rutina pentru evidentierea acestor celule. Cu hematoxilina-eozina celula lui Langerhans apare ca o celula dendritica, putandu-se confunda cu melanocitul. De altfel mult timp celulele lui Langerhans au fost considerate, eronat, ca fiind melanocite epuizate, pe cale de eliminare. Spre deosebire de melanocite insa, acestea sunt aurofile si dopa-negative. Totusi cu clorura de aur, care a permis lui LANGERHANS in 1861 sa descopere aceste celule da rezultate inconstante. Alte coloratii ( cu iodura de osmiu sau albstru de metil) nu sunt specifice, colorand si melanocitele. Parafenilendiamina si dopamina au o specificitate buna, dar necesita tehnici laborioase. In ultimii ani s-au pus la punct tehnici histoenzimatice, dintre care tehnica de evidentiere a adenozintrifosfatazei este specifica si permite obtinerea unei bune imagini a celulelor lui Langerhans.
Totusi, SCHLEGER si BEAN constata ca o trasatura aparent unica pentru celulele lui Langerhans din pielea bovinelor, negativitatea reactiei pentru adenozintrifosfataza utilizand pentru identificarea acestora reactia pentru fosfataza alcalina. Specificitatea acestor reactii este insa discutata.
La microscopul electronic celulele lui Langerhans apar lipsite de tonofilamente si desmozomi. Au un nucleu lobulat iar citoplasma contine o zona Golgi bine marcata, reticul endoplasmatic dezvoltat, lizozomi, vacuole, picaturi de lipide, glicogen, melanozomi fagaocitati. In toata citoplasma, dar cu precadere in vecinatatea aparatului Golgi si a membranei citoplasmatice, contin niste granule considerate markeri ultrastructurali specifici. Acestea imbraca aspectul unor bastonase sau rachete, formate din organite emisferice cu structura membranoasa. Membrana lor periferica se dilata de obicei la o extremitate, pentru a forma o vezicula.
S-au descris doua tipuri morfologice de celule Langerhans:
celule de tip I, asezate deasupra stratului bazal al epidermului, care au citoplasma foarte clara, cu numeroase dendrite, granule Langerhans si care dau o reactie adenozintrifosfatazica intensa;
celule de tip II, localizate in stratul bazal, care au citoplasma densa, slab dendritica si sunt sarace in granule specifice, fiind considerate celule mezenchimale pe cale de diferentiere.
MASUTAMI a aratat ca la soarece celulele prelangerhansiene nedeterminate dobandesc, sub actiunea sensibilizarii de contact, granule Langerhans.
Investigatiile imunologice au permis identificarea la nivelul celulelor Langerhans a antigenilor din grupul complexului major de histocompatibilitate (CMH) clasele I si II-a, a antigenilor notati CD1 si considerati cei mai buni markeri ai limfocitelor T helper.De asemenea s-au identificat receptori pentru regiunea Fc a imunoglobulinelor G si pentru fractiunea C3b a complementului.
Celulele lui Langerhans sunt capabile sa capteze antigenii exogeni patrunsi in epiderm ca haptene sau antigeni infectiosi, sa-i prelucreze si sa-i prezinte limfocitelor T. Aceasta functie de prezentare a antigenilor este dependenta de CMH clasa II-a care sunt asociati antigenului in momentul cooperarii celulelor Langerhans cu limfocitele T helper. Acesta constituie primul semnal al activarii limfocitelor T.
Un al doilea semnal este, deasemenea, indispensabil activarii limfocitelor T. Acesta este realizat de interleukina 1 (IL1), secretata adesea de macrofage. La nivelul epidermului producatorul de IL1 poate induce producerea de substante amiloide de catre hepatocite si fibroblaste ca si chimiotactismul pentru neutrofile.
Limfocitele T se gasesc, la nivel cutanat, in special sub forma de populatii celulare situate in stratul papilar al dermului si avand rol analog populatiilor limfoide din corionul mucoaselor impreuna constituind asa-numitul tesut limfoid asociat mucoaselor si pielii.
La animalul cu pielea intacta, in momentul patrunderii unui antigen prin piele, acesta este intampinat de celulele Langerhans, care il vor prelucra si il vor prezenta limfocitului T helper, care la randul lui va stimula limfocitul capabil sa produca raspuns specific impotriva antigenului respectiv. Cea mai importanta este activarea limfocitului B, care va evolua spre stadiul de plasmocit, capabil sa produca imunoglobuline. In functie de particularitatile antigenului, limfocitul T helper mai poate determina activarea limfocitelor Tc (citotoxice), implicate in distrugerea celulelor infectate de particule virale si in reactia de respingere a grefelor sau activarea celulelor NK ( natural killer) cu rol in distrugerea celulelor tumorale.
Limfocitele T conectate de antigen mai primesc un al doilea semnal prin IL1 produsa de cheratinocite si celulele Langerhans si raspund acestui semnal prin secretia de interleukina 2 IL2. Din acest moment, limfocitele astfel sensibilizate vor migra cu 'imaginea' antigenului in organele limfoide, unde vor prolifera intens generand o clona celulara capabila sa raspunda promt antigenului care a declansat formarea ei si care se va raspandi in intreg organismul.
Experientele pe soareci iradiati cu ultraviolete, la care celulele lui Langerhans sunt scoase din functie, au sugerat existenta la nivelul epidermului si a unui al doilea tip de celule prezentatoare de antigen, rezistente la radiatiile ultraviolete.
Aceste celule au fost descoperite de catre GRANSTEIN in 1984. Ele sunt tot celule dendritice, ca si celulele lui Langerhans, dar spre deosebire de acestea, ele prezinta antigenul limfocitelor T supresoare. Spre deosebire de limfocitele T helper, care recunosc antigenul cu ajutorul CMH clasa I. Limfocitul T supresor intervine in modularea raspunsului imun, limitand raspunsul fata de un antigen dat, atat in reactiile mediate umoral (prin exprimarea transformarii limfocitului B in plasmocit), cat si in cele mediate celular. Acestea ar explica reactivitatea remanenta (si trasferabila) a soarecilor iradiati cu ultraviolete la dinitrofluorbenzen.
In cazul unor traumatisme cutanate minore si repetate, insotite de smulgerea epidermului ca si la animalele iradiate cu ultraviolete, antigenii vor lua contact cu celulele Granstein.
Exista si antigeni care prin proprietatile lor intrinseci prefera contactul cu celulele Granstein.
In procesele de aparare cutanata, un rol important revine mastocitelor ('pazitorii portilor de intrare'). Membrana celulara a acestora prezinta receptori pentru IgE si numeroase prelungiri fine, utilizate pentru incorporarea materialului brut, necesar sintezei de mediatori chimici. Mediatorii chimici stocati in granulele mastocitare sunt reprezentati de: histamina, serotonina, substanta de atragere a eozinofilelor din procesele anafilactice (EAS-A sau ECF-A) si arilsulfataza, enzima care distruge leucotrienele C4 sau substante de reactie lenta a anafilaxiei. Diversi factori patogeni si in mod special, antigenii capabili sa cupleze IgE, determina degranularea mastocitelor (eliberarea mediatorilor), cu efect asupra microcirculatiei. Un alt efect notabil este chemotactismul pentru eozinofile. Acestea intervin in procesele alergice, avand o afinitate deosebita pentru IgE (reagine) si pe de alta parte, au un potential antihistaminic ridicat, observandu-se in teritoriile bogate in acest mediator. In plus, este bine cunoscuta eozinofilia in bolile parazitare.
Fata de reactiile la antigenii exogeni, cele in care sunt implicati antigenii endogeni au un caracter particular. Astfel, in bolile autoimune din grupul penfigus la suprafata cheratinocitelor se elaboreaza anumiti antigeni de natura glicoproteica, fata de care organismul imun va reactiona. Autoanticorpii se leaga de antigenii de suprafata si determina sinteza activatorului de plasminogen. Cresterea locala a plasmei determina pierderea coeziunii celulelor, prin proteoliza rezultand vezicule si bule intermediare.
Tegumentul raspunde prin reactii imune nu numai la alergenii patrunsi pe cale cutanata ci si la cei patrunsi in organism pe cale digestiva, respiratorie, etc. Mucoasa digestiva este in contact cvasipermanent cu alergenii, constituiti din proteine alimentare sau molecule de masa mica susceptibile de a se comporta ca haptene. Majoritatea acestor proteine sunt hidrolizate de catre enzimele proteolitice digestive. O parte sunt complexate de IgA. Totusi o mica parte din aceste proteine alergice pot patrunde mucoasa intestinala intacta, determinand producerea de anticorpi circulanti. Pasajul proteinelor alergice prin mucoasa intestinala este favorizat de leziunile inflamatorii sau degenerative ale acesteia. Pe de alta parte, insuficienta hepatica are drept consecinta insuficienta degradare a proteinelor cu potential alergizant sau a altor molecule care pot juca rolul de haptene.
Din cele prezentate reiese faptul ca in realizarea rolului protector al pielii, in special fata de agentii biotici, o importanta deosebita o detine functia imunitara a acesteia. Legaturile ultrastructurale, biochimice si imunologice dintre timus si piele ca si existenta tesutului limfoid asociat pielii vin sa justifice considerarea pielii ca organ imunitar.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2914
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2026 . All rights reserved