| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
Reactiile de hipersensibilitate
La omul normal, RI (Raspunsul Imun) Umoral si Celular prezinta doua particularitati fundamentale:
RIU/C este un mecanism de protectie care asigura doua tipuri de aparari naturale:
apararea antibacteriana, antivirala si antiparazitara;
supravegherea si apararea antitumorala si anticanceroasa.
RIU/C nu se insoteste nicicand de efecte defavorabile pentru organism deoarece este perfect adaptat calitativ si cantitativ la natura si intensitatea stimulului antigenic.
I. RIU este normal daca raspunsul declansat fata de antigen se face printr-o clasa adecvata de imunoglobuline: IgM, IgG sau IgA. Acest criteriu nu este indeplinit in cazul hipersensibilitatii de tip I (IgE).
II. RIU este normal daca Ig elaborate nu ating concentratii exagerat de mari in raport cu cantitatea necesara pentru neutralizarea si indepartarea antigenului declansator. Acest criteriu nu se indeplineste in toate hipersensibilitatile umorale si in majoritatea bolilor autoimune.
III.RIU este normal daca nu se elaboreaza cantitati crescute de complexe imune circulante. Acest criteriu nu se indeplineste in hipersensibilitatea de tip III.
I. Antigenele declansatoare nu sunt expuse pe membranele celulelor somatice non-APC;
II. Celulele imunocompetente nu sunt colonizate de microorganisme cu habitat intracelular.
Ambele criterii de normalitate a RIC nu sunt indeplinite in hipersensibilitatea de tip IV.
I. Strict localizate, adica daca se desfasoara la poarta de intrare a antigenului in organism.
II. Strict controlate, adica daca exista un echilibru functional fin intre functia limfocitelor Th si functia limfocitelor Ts.
III. Neinsotite in evolutia lor de aparitia unor leziuni tisulare; daca acestea apar, sa fie de minima importanta si sa nu persiste in timp.
Definitie: Reactiile de hipersensibilitate sunt RI anormale, patologice, lipsite total de efectul de protectie, ele evoluand cu aparitia unor leziuni tisulare severe ca urmare a declansarii unor procese inflamatorii acute sau cronice.
Toate reactiile de hipersensibilitate sunt reactii inflamatorii patologice.
HS I se datoreaza sintezei unor clase neadecvate de anticorpi, IgE.
HS II se datoreaza prezentei anormale a antigenelor pe membranele celulelor self, antigene fata de care se elaboreaza anticorpi citotoxici din clasele IgM si IgG.
HS III se datoreaza elaborarii unor cantitati exagerate de CIC (Complexe Imune Circulante).
Tipurile I, II, III se numesc reactii de hipersensibilitate de tip imediat pentru ca manifestarile clinice se instaleaza la putin timp dupa contactul cu antigenul declansator, deoarece aceste HS au la baza o mediatie umorala.
HS IV mai este numita hipersensibilitatea de tip intarziat pentru ca manifestarile clinice caracteristice apar la distanta (in timp) fata de contactul antigenic declansator si persista mult, avand la baza o mediatie celulara.
Toate cele patru tipuri de HS se insotesc in mod obligatoriu de aparitia unor leziuni tisulare severe care au la baza lor procese inflamatorii:
acute, in cazul HS imediate;
cronice, in cazul HS de tip intarziat.
Definitie: Hipersensibilitatea de tip I este un RIU anormal declansat la contactul organismului cu antigene banale/obisnuite, raspandite in mediu, prezente in alimente sau in structura medicamentelor, antigene fata de care populatia generala nu reactioneaza; HS I se datoreaza unei hiperproductii de IgE care este conditionata genetic.
Bolnavii cu HS I prezinta doua trasaturi:
Manifestari de tip alergic (un RI deviat ce consta intr-o reactie la antigene fata de care populatia generala nu prezinta un raspuns clinic manifest).
Aceste antigene declansatoare ale HS I se numesc alergene. Alergenele sunt antigene complete timodependente si au imunogenitate redusa. Sunt foarte raspandite in mediu, contactul cu ele fiind obligatoriu pentru intreaga populatie.
Exemple de alergene: polenuri, praful de casa (acarienii microscopici).
Bolnavii prezinta un teren atopic ce consta in hiperproductie de IgE, conditionata genetic.
Schema de principiu a HS I cuprinde 2 etape:
Etapa contactului antigenic inductor: are loc cu ocazia primului contact cu antigenul declansator al HS I si este asimptomatica din punct de vedere clinic.
Etapa contactului antigenic declansator - are loc cu ocazia unui contact antigenic ulterior (contactul numarul 2, 3, ., n) cu antigenul declansator.
Exemple: rinita alergica, astmul bronsic extrinsec, gastroenterita alergica (dermatita atopica).
Ambele etape sunt rezultatul contactului dintre organism si alergenele care penetreaza barierele dintre mediu si organism:
tesutul cutanat;
mucoasa bronsica;
mucoasa gastrointestinala.
Antigenele patrund usor in organism si activeaza niste RIU deviate (patologice) adica hiperproductie de Ig clasa IgE. Aceste IgE sunt anticorpi citofili (nu raman liberi) si se ataseaza de membranele celulare ale mastocitelor/bazofilelor din intreg organismul. Fixarea lor pe suprafata membranelor se face prin receptorii specifici FceR.
La finele etapei de inductie are loc hiperproductie IgE cu tapetarea tuturor mastocitelor si bazofilelor din organism.
Alergenul este recunoscut de IgE care sunt deja atasate pe suprafetele mastocitelor, bazofilelor. Apar anumiti stimuli activatori ce se transmit in profunzimea mastocitelor, bazofilelor. Activarea acestor celule este urmata de degranularea lor si eliberarea unor cantitati mari de:
histamina;
PAF;
SRSA;
Pg, LT, Tx A2.
Toate aceste substante sunt mediatori ai inflamatiei acute.
In functie de locul eliberarii acestor mediatori apar diverse manifestari clinice:
de tip rinita alergica daca agentul alergen penetreaza mucoasa nazala, iar mediatorii sunt eliberati la nivelul acesteia;
astm bronsic daca antigenele patrund in mucoasa bronsica iar mediatorii sunt eliberati in aceasta mucoasa;
dermatita atopica daca antigenele patrund prin tesutul cutanat iar mediatorii sunt eliberati la locul de patrundere.
Din punctul de vedere al elementelor participante la HS I:
Alergenul este un element normal si cu larga raspandire;
Macrofagele si bazofilele sunt elemente normale;
Anticorpii de tip IgE sunt patologici, atat prin cantitatea mare in care sunt produsi cat si pentru motivul ca fac parte dintr-o clasa neadecvata.
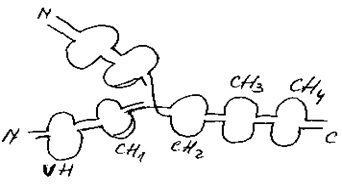
Structura IgE: IgE sunt tetrameri H2L2. Lantul usor L are doua domenii iar lantul greu H are 5 domenii (1 variabil si 4 constante).
Figura
Concentratia IgE este in mod fiziologic mica, de ordinul nanogramelor la ml (1 UIs 2,5 ng/mL), dar variaza foarte mult in functie de varsta:
la nastere: 0.2 UI;
la 12.13 ani: 200 UI (concentratie maxima);
la adult: 30.50 UI.
Durata de viata a IgE este foarte redusa (2.3 zile).
Particularitati functionale ale IgE:
IgE nu traverseaza placenta;
IgE nu activeaza complementul;
IgE sunt anticorpi citofili, fixarea la membrana celulara facandu-se prin receptorii specifici FceR.
FceR cad in doua categorii:
A. FceR1:
sunt receptori de inalta afinitate pentru IgE;
sunt distribuiti restrans in organism, fiind exprimati doar pe membranele macrofagelor si bazofilelor;
sunt formati din trei tipuri de lanturi:
q Lantul alfa este orientat cu ambele capete intracelular; este receptorul propriu-zis pentru IgE, angajand legaturi cu domeniile CH1 si CH2 ale IgE;
q Lantul beta are de asemenea ambele capete orientate intracelular; are numai rol structural;
q Lanturile gama, in numar de doua sunt transmembranare, orientate cu capatul carboxiterminal intracelular iar cu cel aminoterminal, extracelular; in cazul in care IgE atasate de membrana recunosc un antigen, lanturile gama transmit semnale activatoare. In momentul legarii antigenului Fab al IgE sufera transformari conformationale care se repercuteaza si la nivelul lanturilor alfa. Mai departe semnalele activatoare sunt transmise prin unele sisteme tip mesager secund ale macrofagelor / bazofilelor a caror activare o determina.
B. FceR2 (CD23)
sunt receptori de joasa afinitate pentru IgE;
sunt foarte larg raspanditi pe membranele celulelor sanguine: monocite, macrofage, eozinofile, trombocite si pe suprafata unor limfocite B (limfocitele B e sunt CD23S);
sunt formati dintr-un singur lant polipeptidic foarte lung, orientat contrar lantului alfa al FceR1, cu capatul carboxiterminal extracelular si capatul aminoterminal, intracelular;
in structura lantului FceR2 intra trei segmente diferite (de la capatul extracelular la cel aminoterminal):
q segmentul lectinic este receptorul propriu-zis pentru IgE; legatura cu IgE se realizeaza prin intermediul unor carbohidrati bogat reprezentati la nivelul CH3 si CH4 in IgE;
q segmentul de clivaj poate fi clivat in prezenta anumitor enzime proteolitice eliberate dupa activarea celulei;
q segmentul transmembranar.
FceR2 (CD23) poate exista in doua forme:
Forma atasata de membrana functioneaza ca receptor IgE;
Forma libera (solubilizata) rezulta in urma clivarii FceR2 in zona de clivaj, sub actiunea unor enzime proteolitice eliberate dupa activarea celulei; este pusa in libertate portiunea lectinica a FceR2 (situata spre capatul carboxiterminal) care se noteaza CD23s (solubilizat); CD23s pierde capacitatea de fixare a IgE si capata functii de interleukina. Alaturi de IL-4, CD23s stimuleaza productia de IgE, ambele activand mecanismul de comutare izotipica spre IgE.
Reglarea izotipica a sintezei IgE: la sinteza IgE participa trei categorii de celule:
APC: macrofage;
Limfocite cu rol imunoreglator: limfocite Th2e (coopereaza in egala masura si simultan cu limfocitele Bm (obisnuite) care prezinta pe suprafata lor un BCR tip IgM si cu limfocitele Be care exprima pe suprafata lor CD23)
limfocitele Be
limfocitele Ts antigen specifice;
limfocitele Th1 (in numar redus).
Limfocitele Bm sunt celule efectorii.
Intre aceste celule are loc o cooperare ce parcurge mai multe etape:
Antigenele sunt preluate de macrofage, care le prelucreaza si le exprima pe suprafata lor membranara in complex cu MHC I/II, prezentandu-le catre toate cele 4 tipuri de limfocite imunoreglatorii. Limfocitele Th2e sunt primele activate si incep sa coopereze imediat cu limfocitele Bm (cooperare noncognitiva prin IL-4) si limfocitele Be (tot prin IL-4).
Primul stimulat este limfocitul Be ce isi elibereaza receptorul de suprafata CD23 ce devine CD23s (solubilizat). CD23s impreuna cu IL-4 activeaza limfocitul Bm, stimuland la nivelul acestuia productia de IgE ca urmare a activarii mecanismului de comutare izotipica.
Functia stimulatoare a CD23s depinde foarte mult de starea sa biochimica; glicozilarea se realizeaza extemporaneu chiar in cursul cooperarii celulare si este sub controlul a doi factori:
GEF (factorul de activare a glicozilarii), produs doar de subsetul limfocitelor The
GIF (factorul de inhibare a glicozilarii), produs de limfocitele Ts.
Actiunea GEF predomina asupra actiunii GIF si in consecinta se formeaza CD23s glicozilat care se ataseaza de suprafata limfocitelor Bm la receptorul CR2 (CD21, al 2-lea receptor pentru complement). Semnalele declansate activeaza mecanismul de comutare izotipica, care inhiba productia de IgM si favorizeaza productia de IgE. La controlul sintezei IgE mai participa si limfocitele Th1.
La formarea limfocitelor Th2 se diferentiaza si un numar mic de limfocite Th1; acestea (Th1) controleaza productia de IgE prin intermediul interferonului gama care se opune mecanismului de comutare izotipica. In mod fiziologic, productia de IgE este foarte bine controlata si foarte redusa, existand factori favorizanti (GEF si IL-4) si factori de inhibitie (IFN-g si GIF) ai productiei IgE. Echilibrul realizat prin intermediul acestor factori este foarte fin.
Mecanismele hiperproductiei de IgE la pacientii cu boli alergice
Cresterea (dublarea) numarului de limfocite Be duce la cresterea sintezei IgE deoarece limfocitele Be sunt sursa de CD23;
Exista o triplare a numarului limfocitelor Th2e care stimuleaza prin IL-4 productia de IgE, iar prin GEF favorizeaza glicozilarea CD23s, fenomene care duc la cresterea productiei IgE.
Scaderea numarului si functiei limfocitelor Ts la bolnavii cu boli alergice: prin carenta de GIF are loc o glicozilare excesiva a CD23s.
Scaderea numarului si activitatii limfocitelor Th1 este insotita si de deficiente in apararea celulara (carenta de IFN-g
Expresia pe membrana APC a unor molecule MHC II suprastimulante pentru limfocitele Th2 sau a unor molecule MHC I substimulante pentru limfocitele Ts.
In cazul limfocitelor Th2, urmatoarele varietati MHC II sunt suprastimulante:
HLA-DR2;
HLA-DR3;
HLA-DR5.
In cazul limfocitelor Ts, urmatoarele varietati MHC I realizeaza o slaba prezentare a antigenului: HLA-DR8.
In cazul in care pacientul exprima una din aceste varietati MHC, se creaza un dezechilibru functional intre limfocitele Th2 si Ts, manifestat prin hiperproductie de IgE. IgE se depoziteaza pe suprafata macrofagelor si a bazofilelor. Ca urmare a recunoasterii antigenice IgE depozitate pe suprafetele acestor celule duc la activarea lor cu eliberare de mediatori.
Mecanismele activarii macrofagelor/ bazofilelor
Macrofagele si bazofilele sunt celulele efectorii ale hipersensibilitatii de tip I. Activarea lor survine numai dupa ce a avut loc recunoasterea antigenica pe suprafata lor. Receptorii FceR1 de pe suprafata acestor celule fixeaza multe macromolecule IgE si atunci cand are loc recunoasterea alergenelor de catre IgE, activeaza celulele.
Stimulii activatori se transmit prin lantul gama (n.ed. please verify gamma chain 'cause I'm not sure) spre trei sisteme tip mesager secund: sistemul adenilat-ciclaza (AC), proteinlipaza C (PLC) si proteinlipaza A2 (PLA2).
Prin adenilat ciclaza cresc concentratia AMPc (adenozin-monofosfat ciclic) si activitatea PKA (proteinkinaza A).
Prin sistemul PLC fosfatidilinozitolul membranar este desfacut in IP3 (inozitoltrifosfat) si DAG (diacilglicerol). IP3 provoaca o crestere a concentratiei calciului (hipercalcitie) iar DAG induce o crestere a activitatii PKC (proteinkinaza C).
Enzimele PKA, PKC si hipercalcitia conduc la activarea sistemului de microtubuli si microfilamente din citoplasma celulelor efectorii ale HS I, fenomen care are drept rezultat expulzia granulatiilor. Prin degranulare se pun in libertate mediatori preformati.
Prin sistemul PLA2 se genereaza PAF (n. ed. Platelet Activation Factor; factorul de activare plachetar) care ramane ca atare si acid arahidonic care poate urma doua cai metabolice:
pe calea COX (ciclooxigenaza) este desfacut in Pg (prostaglandine) si TxA2 (tromboxan A2);
pe calea LOX (lipooxigenaza) serveste la sinteza de LT (leucotriene).
Hy (histamina) are efecte:
vasomotorii: vasodilatatie si cresterea permeabilitatii capilare→ edem;
bronhomotorii: bronhoconstrictie, edem al mucoasei bronsice, hipersecretie de mucus;
spasmogene intestinale: contractia musculaturii netede intestinale si edem al mucoasei intestinale;
hemodinamice generale: Hy se elibereaza direct in circulatie si poate provoca socuri anafilactice (colaps) prin faptul ca provoaca o hipotensiune prin vasoplegie generalizata; deasemenea deprima activitatea de pompa a cordului, implicand instalarea aritmiilor cu ritm rapid (> 160.180/min).
Mediatorii chemotactici sunt:
ECFA (factor chemotactic pentru eozinofile al anafilaxiei);
NCFA (factor chemotactic pentru neutrofile al anafilaxiei) este echivalentul IL-8;
Unele enzime (proteaze: chimaze, triptaze).
PAF (factorul activator plachetar) are efecte:
bronhomotorii: bronhoconstrictie;
spasmogene intestinale;
vasomotorii: vasodilatatie si cresterea permeabilitatii membranei capilare.
deprimante cardiace: bradicardie accentuata (35.40/min) si scaderea debitului coronarian (coronaroconstrictie);
stimuleaza agregarea plachetelor;
activeaza leucocitele.
Derivati ciclooxigenati ai acidului arahidonic (AAH):
PgD2 - efect vasomotor si bronhomotor;
PgF2α - efecte bronhomotorii;
TxA2 - efect bronhomotor si de stimulare a agregarii plachetare.
Derivati lipooxigenati ai acidului arahidonic:
SRSA este un amestec de trei leucotriene (C4, D4 si E4) cu urmatoarele efecte:
q bronhomotor;
q vasomotor;
q inotrop negativ.
LT B4 este un chemoatractant foarte activ pentru PMN; are si efect bronhomotor.
Sindromul clinic se caracterizeaza prin aparitia unor crize de dispnee paroxistica, cu caracter expirator, care au loc la contactul dintre alergen si mucoasa bronsica.
Crizele se datoreaza bronhoconstrictiei, edemului mucoasei bronsice si hipersecretiei de mucus si prezinta unele particularitati in functie de stadiul evolutiv al bolii.
In stadiul initial, crizele sunt scurte, fara mare intensitate; intre crize bolnavul este aparent normal;
In stadiul intermediar, crizele sunt mai frecvente si mai intense; intre crize apar semne de hiperreactivitate bronsica;
In stadiul terminal al astmului bronsic, crizele sunt frecvente si prelungite iar intre ele apar semne paraclinice si clinice de disfunctie ventilatorie obstructiva (DVO).
Crizele de astm sunt determinate de eliberarea in submucoasa bronsica a mediatorilor bronhospastici si chemoatractanti, dupa ce alergenele au fost recunoscute.
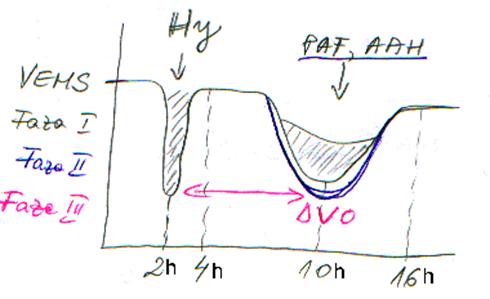
Mediatorii bronhospastici altereaza tonusul vasomotor. La nivelul miocitelor bronsice toti acesti mediatori activeaza sistemul cGMP, declansand bronhoconstrictia. Bronhospasmul este multiplu, determinat de doua categorii de mediatori (primari si secundari) iar la examenul spirografic se evidentiaza prin scaderea VEMS (volum expirator maxim pe secunda).
In timpul crizei apar doua tipuri de bronhospasm:
Apare la cel mult 2 ore dupa debutul crizei, la circa 4 ore tonusul revine la normal. Acest bronhospasm este initiat de histamina; efectul este limitat in timp datorita actiunii enzimelor histaminaza si N-metiltransferaza, care distrug histamina.
Apare la maxim 10 ore, tonusul revine la normal dupa circa 16 ore de la debut. Acest al 2-lea bronhospasm este determinat de mediatorii neoformati si de aceea se instaleaza mai lent si este mai persistent.
Figura
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2064
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2026 . All rights reserved