| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
Definitie: orice hipersensibilitate imediata se datoreaza factorului antigenic declansat pe membrana celulei somatice non-APC, antigen fata de care se elibereaza Ac citotoxici, urmand distrugerea celulei din tesutul tinta: reactie citotoxica.
Etape:
Declansarea cascadei complementului pe calea clasica
Generarea locala a unui proces inflamator acut
ADCC (citotoxicitate celulara anticorp-dependenta)
Pe membrana se afla: Ag atasate (mai rar) si constitutive(frecvent), se mai afla receptorii pentru Ac IgG, deci se formeaza complexe imune. Mai exista pe membrana si C1q care este recunoscut de complexele Ag-Ac pe membrana, astfel fixeaza C1r si C1s si se activeaza cascada complementului; C3 convertaza actioneaza asupra C3, rezulta C3a si C3b. C3b se depoziteaza pe membrana si opsonizeaza. Se depun de o parte si de alta a complexelor imune.
C3b este recunoscut de macrofagele intratisulare si debuteaza activarea acestora, astfel C5 convertaza actioneaza asupra lui C5 si il cliveaza in C5a si C5b, care formeaza MAC (vezi cursul de complement), se insera in membrana celulelor tinta si distrug celula prin liza osmotica.
Nu toate celulele pe suprafata carora se activeaza complementul pot fi distruse per prima, astfel se deosebesc doua categorii de celule:
celule anucleate ce nu rezista activarii complementului, pot fi distruse per prima, nu dispun de mecanisme reparatorii ale membranei (eritrocite, trombocite).
celule nucleate, ce rezista la actiunea distrugatoare a complementului deoarece dispun de mecanisme reparatorii ale membranei (au sinteza proteica crescuta)
Procesele inflamatorii acute favorizeaza net distrugerea celulelor nucleate, care este declansata ca urmare a eliberarii anafilatoxinelor (C3a, C5a). Odata eliberate stimuleaza mastocitele si bazofilele, acestea dispun de mecanisme speciale pentru C3a si C5a si elibereaza mediatori preformati (mai ales histamina), ulterior ajungand si la mediatori de neoformatie(de novo).
Au actiune vasomotorie: histamina este vasodilatator, creste permeabilitatea vasculara, are actiune de scurta durata. Vasodilatatia este mentinuta si accentuata si prin eliberarea de CK si IL produse de macrofagele ce s-au activat in etapa anterioara: IL1(forma solubila), TNF actioneaza pe celulele endoteliale locale, crescand intens activitatea ciclooxigenazei endoteliale, se face productie crescuta de PG2 si prostacicline (cu efecte vasodilatatoare de lunga durata).
PAF si LT au si ele efecte vasodilatatoare.
Vasodilatatoarele aduc un flux mare de celule ce adera (mai ales leucocitele) la endoteliul vascular.
IL si TNF favorizeaza foarte mult aderarea celulelor la endoteliu, cresc expresia de LFA, pe membranele leucocitelor, si de ICAM pe membranele celulelor endoteliale.
Celulele trec prin diapedeza in tesutul antigenic, datorita unor factori chemoatractanti: C3a si C5a cu efect chemoatractant pe monocite si neutrofile, LT B4 cu efect chemoatractant pentru leucocite, IL1 cu efect pe celulele K. Astfel se constituie un infiltrat celular proinflamator. La contactul neutrofilelor, celulelor Killer, macrofagelor cu celulele tinta se declanseaza etapa a treia.
Celulele proinflamatorii distrug celulele tinta parcurgand cateva etape:
Recunosc tintele celulare prin recunoastere imunologica nespecifica, diferita de la o celula la alta.
q Neutrofilele: dupa complexele imune existente pe suprafata lor, ele dispun de receptori FCgR
q Celulele Killer: dupa complexele imune, au pe suprafata lor CD16 (FCgR3)
q Macrofagele: prin recunoasterea complexelor imune (FCgR) si prin contact cu celulele tinta prin C3b depozitat pe membrana si receptorii pentru complexul CR
Celulele efectorii se activeaza metabolic si elaboreaza unii mediatori citotoxici prin care distrug tintele celulare.
Neutrofilele se degranuleaza si elibereaza enzime, ele mai produc cantitati crescute de radicali liberi de oxigen ce distrug membranele celulelor tinta. Celulele Killer elibereaza perforina. Macrofagele se degranuleaza si elibereaza enzime si secreta cantitati mari de radicali liberi de oxigen.
Cele trei etape sunt parcurse integral sau nu in functie de locul reactiilor:
intratisular: parcurg integral etapele
intracirculator: doar etapele 1 si 3
Anticorpii de tip M sau G citotoxici de obicei pot declansa 2 mecanisme cu potential citotoxic: pot declansa cascada complementului sau pot media desfasurarea ADCC.
Sunt rezultatul unor RIU de tip imunizari, care, in functie de originea Ag, sunt de doua tipuri cu doua mecanisme de generare a Ac:
Alloimunizare: dau Ac citotoxici datorita declansarii RIU fata de celule provenite din tesuturi straine, de doua categorii:
a) celule din tesuturi allogene: apartin unor indivizi diferiti ce fac parte din aceeasi specie; sunt celule non-self doar ca exprima pe suprafata lor haplotipuri diferite de MHC.
b) celule xenogenice ce apartin unor indivizi diferiti ce fac parte din specii diferite.
Exemplu alloimunizare: posttranfuzionala, fetomaterna, posttransplant.
Autoimunizarea: Ac citotoxici ce apar datorita unui RIU fata de propriile tesuturi, secundari ruperii tolerantei imune fata de celulele self.
Exemple: anemie hemolitica autoimuna, LED, scleroza in placi, miastenia gravis.
Se datoreaza transferului prin transfuzii de sange total a unei mase eritrocitare de grup sanguin incompatibil. Eritrocitele donor au alte specificitati Ag decat cele ale primitorului.
Importanta grupei sanguine:
ABO dau majoritatea reactiilor posttransfuzionale, este o grupa completa ce este formata din Ag eritrocitar specific pentru fiecare grupa, si Ac specifici sau aglutinine.
Ag de grup ABO sunt formate din carbohidrat de baza ce intra in structura tuturor Ag ABO, un zaharid terminal, prin care Ag difera intre ele.
Insertia zaharidului terminal se face prin activarea a trei glicozil transferaze (codate pe cromozomul 9); se activeaza seriat in procesul de maturare a Ag ABO ce se termina in perioada neonatala si incepe in perioada embriofetala.
In perioada embriofetala eritrocitele prezinta pe membrana Ag I (initial) format din patru carbohidrate:
Figura reprezinta carbohidratul de baza
NAGA = N-acetil-galactozamina
![]()
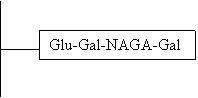
Ag I este o structura total non-imunogena
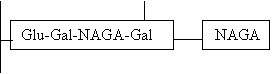
O foarte mare parte din ele (mai putin de 5%) raman nemodificate in procesul de maturare, iar 95% sufera modificari succesive sub actiunea glicozil-transferazei spre varful perioadei fetale. Mai intra in actiune si gena H de pe cromozomul 9 si se activeaza fucozil-transferaza ce insera la galactoza terminala un rest de fucoza, Ag H caracteristic pentru grupul sanguin O (45%).
Ag H este structura non-imunogena.
La cei cu alte grupe de sange, Ag H sufera alte modificari:
Prin intrarea in functie a genei A (cz. 9), se activeaza galactozamin transferaza si se insera la galactoza terminala inca un rest NAGA si da grupul sanguin A. Ag A este o structura intens imunogena (40% din populatie).
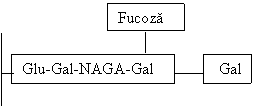
Fucoza
![]()
Prin intrarea in functie a genei B se transfera un rest galactozil (prin activarea galactozil transferazei codata de cz 9), formandu-se grupa B (10% din populatie), cu Ag B ca structura intens imunogena.
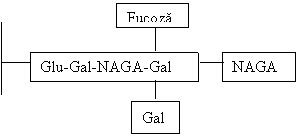
![]()
Pentru grupa AB se activeaza atat gena A cat si gena B si se vor alipi la Gal terminala atat restul de NAGA cat si cel de Gal, cu frecventa de mai putin de 5% din populatie.
![]()
Distributia Ag ABO este larga in organism, sunt bine reprezentate pe suprafata eritrocitelor, sunt si pe membrana leucocitelor, plachetelor, mari majoritati a celulelor somatice (rinichi, tub digestiv, plaman, cord) .
Pentru practica transplantului si a transfuziilor sunt importante aglutininele, Ac de un anumit tip, elaborati pentru a face fata tolerantei fata de propriile eritrocite si non-tolerantei fata de cele non-self.
La organismul normal nu se sintetizeaza Ac fata de propriile Ag
La organismul normal se elaboreaza Ac numai fata de grupul sanguin absent sau nemodificat genetic:
Ag A prezinta Ac anti B
Ag B prezinta Ac anti A
Ag H prezinta Ac anti A anti B
Ag AB nu are anticorpi
Schema tolerabilitatii intre cele 4 grupe sanguine:
 Grupa
A
Grupa
A
Ac α B
Ag A
![]()
 Grupa 0 Grupa
AB
Grupa 0 Grupa
AB
Ac α A Ag A
Ac α B Ag B
Grupa B
Ac α A
Ag B
Tolerabilitatea este relativa.
Exemplu: transfuzie sange de grupa O la grupa A: se transfera eritrocitele si Ac anti B si anti A, sunt bine primite, dar sunt mari imunogene, daca vor recunoaste Ag A dau o reactie citotoxica si imunoliza.
La organismul normal riscul de imunoliza este redus deoarece:
Titrul de Ac α A sau α B este foarte redus
Ac α A sau α B in organismul primitor scade si mai mult
parte din Ac sunt deviati de la contactul cu hematiile fiind cuplati cu Ag de pe celulele somatice
Riscul devine maxim cand se doneaza sange de la donatori periculosi: politransfuzati, gravide.
Gravida: are grupa O si Ag H, Ac α A, Ac α B, iar fatul are grupa A. La nastere trece un mic numar de hematii din circulatia fetala in circulatia materna si da un RIU destul de intens mult dupa nastere, generandu-se Ac α A sintetizati de novo, deci creste concentratia de Ac α A la mama. Daca se face o transfuzie catre un subiect de grupa A, eritrocitele de grup sanguin O sunt foarte bine tolerate, dar plasma are o concentratie crescuta de Ac α A, o mare parte au contact cu Ag A al primitorului si se declanseaza cascada complementului ce va distruge o mare parte din eritrocitele primite.
Anticorpii α A sau α B pot fi naturali sau imuni.
Anticorpii α A/B naturali sunt IgM, sunt prezenti permanent in circulatie si in concentratii mici, nu traverseaza placenta, activeaza cascada complementului sau aglutineaza in special la temperaturi mai joase (se mai numesc si Ac la "rece").
anticorpii α A/B imuni apar numai episodic, ca urmare a unei stimulari Ag cunoscute, sunt din clasa IgG, traverseaza placenta, activeaza cascada complementului, cand exista, se afla mereu in concentratii foarte mari.
Cele doua tipuri de Ac sunt implicate in generarea de reactii citotoxice posttransfuzie, de obicei la subiectii care primesc fie transfuzii compatibile de la donatori periculosi sau transfuzii incompatibile.
Aglutinarea eritrocitelor
ale donorului, daca transfuzia este incompatibila
ale primitorului daca transfuzia este de la donatori periculosi
exista aglutinari preluate de celulele SREE (sistemul reticulo-endocitar) si distruse prin hemoliza extravasculara.
Mentinerea sau agravarea anemiei posthemoragice acute duce la hipoxie tisulara, daca este prelungita duce la aparitia de leziuni minime endoteliale
Activarea cascadei complementului duce la eliberarea in circulatie de anafilotoxine ce stimuleaza difuz macrofagele perivasculare si bazofilele intravasculare, deci se elibereaza in circulatie mediatori pre-neo-formati si duce la vasoplegie generalizata periferica, colaps sever, scaderea circulatiei periferice si hipoxie T se va afecta in primul rand rinichiul prin ischemie renala, apare insuficienta renala acuta, iar daca se mentine duce la necroza paranchimului renal.
Distrugerea eritrocitelor pe suprafata carora s-a activat complementul se face prin hemoliza intravasculara, se elibereaza tromboplastina si se declanseaza coagularea pe cale extrinseca ce duce la CID(coagulare intrinseca diseminata)
Leziunile endoteliale activeaza factorul XII, sa declanseaza coagularea pe cale intrinseca
Macrofagele si bazofilele secreta PAF, TX A2 ce stimuleaza trombocitul si duce la CID prin trombi.
Este declansata de Ag apartinand grupelor Rh. Grupa Rh este o grupa incompleta, are doar un Ag specific de grup Rh. In conditii obisnuite Ac naturali anti Rh nu exista. Daca apar apar numai imuni.
Ag Rh C D E
![]()
![]()
![]()
![]() 5 3
5 3
Sunt sintetizati prin gene de pe cromozomul 1, unde exista 3 gene, dintre cele 3 locusuri predominant este cel al genei D. Exista mai multe posibilitati de recombinare: CDe duce la Ag Rh 1, cDe duce la Ag Rh 2, cde duce la grup Rh -, fara Ag.
Ag Rh 1, Rh 2 se afla strict pe membranele eritrocitare si sunt intens imunogene. Ac anti Rh, cand exista, sunt numai Ac imuni, deci exista pe perioade limitate de timp si in concentratii crescute, au un pronuntat caracter citotoxic si actioneaza asupra unei singure celule tinta: eritrocitul.
La gravidele Rh -, purtatoare de fat cu Rh +:
La travaliu, eritrocitele Rh + trec in circulatia materna, declanseaza post partum un RIU cu elaborare de Ac α Rh, de tip Ig G, in concentratii crescute, ce raman pe perioade lungi de timp si pot trece placenta.
Daca in aceeasi perioada survine o a doua sarcina, Ac trec in organismul fetal si dau reactii Ac-Ag ce duce la hemoliza intravasculara fetala ce va scade foarte mult numarul de eritrocite si va duce la aparitia unei anemii cu hipoxie severa in tesuturile fetale si stimularea secretiei de eritropoetina.
Daca este la varsta mica (fatul), afecteaza hematopoeza extramedulara, daca este la varsta mare afecteaza maduva spinarii; da reticulocitoza, precursori eritroblastici nucleati.
Compensarea nu este eficienta deoarece hematiile sunt foarte slab incarcate cu hemoglobina de tip Hb F, incapabila sa transporte oxigenul si duce la decompensare cardiaca si edem, hidraps fetal si exitus in utero.
Dupa hemoliza duce la hiperbilirubinemie indirecta, nu poate fi conjugata, ramane in circulatie si fiind liposolubila se depune in SNC provocand leziuni ale nucleilor de la baza creierului si icter nuclear.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1768
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2026 . All rights reserved