| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
REACTII IMUNITARE DE HIPERSENSIBILITATE
De cele mai multe ori, in special in procesele infectioase, reactiile imunitare au o finalitate benefica, avand un rol determinant in eliminarea agentilor patogeni. Uneori, dupa contactul cu antigenele (in special moleculare), activarea functiei imunitare are rol prejudiciant, defavorabil asupra organismului, deoarece raspunsul imun se instituie drept cauza si mecanism pentru producerea diferitelor maladii (alergii, boli autoimmune)
Activarea neadecvata a functiei imunitare determina doua categorii de manifestari clinice:
a) starile de hipersensibilitate
b) maladiile autoimune
Diminuarea activitatii sistemului imunitar, genereaza o categorie speciala de manifestari clinice, cunoscute sub denumirea generica de imunodeficiente. Ele pot fi inascute (primare) sau dobandite (secundare).
Starile de hipersensibilitate sunt reactii tertiare, consecutive reactiilor Ag-Ac in vivo. Ele sunt o consecinta a faptului ca procesul de imunizare dupa contactul primar cu antigenul si generarea efectorilor imunitari (anticorpi si limfocite efectoare) nu confera totdeauna o stare favorabila, de rezistenta a organismului. Contactul primar cu antigenul creeaza, uneori, o stare de sensibilizare fata de antigenul respectiv. Sensibilizarea este o stare fiziologica daunatoare organismului si se manifesta, in special dupa contactul organismului cu antigene proteice (din ou, din ser), cu antigenele din polen si mai rar dupa contactul cu antigenele corpusculare (hematii de berbec). La contactul secundar cu antigenul sensibilizant, organismul raspunde prin starile patologice de hipersensibilitate.
Starile de hipersensibilitate sunt consecinta unui raspuns de intensitate prea mare sau a unui raspuns imun neadecvat, care sta la originea leziunilor tisulare. Echivalentul termenului de hipersensibilitate, folosit in mod curent, este cel de alergie (allos, ergon = alta energie).
Termenul de alergie a fost introdus de von Pirquet (1906) si semnifica o reactie imunitara care se exprima cu energie diferita de cea normala, dupa expunerea secundara la un antigen. Ambele denumiri se refera la o reactivitate imunitara de intensitate anormal crescuta, fata de un antigen. In sens stiintific, notiunea de alergie include toate manifestarile care decurg din reactivitatea imunitara, cu o alta energie decat cea fiziologica: reactiile hiperergice, hipoergice si anergice.
In mod curent, alergia se defineste ca o stare de hipersensibilitate, ce rezulta din expunerea la un alergen si se distinge prin supraproductia componentelor imunitare.
Clasificarea starilor de hipersensibilitate
Starile de hipersensibilitate au fost clasificate in raport cu promptitudinea cu care se manifesta:
- reactiile de hipersensibilitate imediata au o dinamica rapida. Se declanseaza in cateva secunde sau minute de la contactul secundar cu alergenul si diminua rapid, in cateva ore, fara semne vizibile, cu exceptia celor foarte grave. Tesutul suport al reactiei este diferit de la o specie la alta. Aproape totdeauna va rezulta distrugerea celulei tinta. Reactiile imediate se desfasoara in tesuturi vascularizate si, de obicei, se manifesta local, dar pot produce si efecte sistemice. Hipersensibilitatea imediata este cea mai raspandita dezordine imunitara la om. Este cea mai frecventa maladie cronica, ce afecteaza circa 25% din populatie in tarile dezvoltate, cu severitate variabila, de la o simpla iritare, pana la periclitarea vietii;
- reactiile de hipersensibilitate subacuta sunt acelea care incep sa se manifeste dupa 1-3 ore de la contactul secundar cu antigenul si inceteaza dupa 10-15 ore. Sunt mediate de IgG sau IgM;
- reactiile de hipersensibilitate intarziata se evidentiaza la 1-2 zile dupa contactul secundar cu alergenul. Persista un interval de cateva zile, pana la cateva saptamani. Aceste reactii sunt mediate de limfocitele T si de macrofage. Nu sunt dependente de factori humorali circulanti si de aceea se pot produce si intr-un tesut nevascularizat. Singura conditie este ca tesutul sa fie situat in apropiere de sistemul vascular, pentru ca limfocitele si macrofagele sa poata migra spre locul unde a fost injectat antigenul.
Reactiile de hipersensibilitate imediata si intarziata se deosebesc prin urmatoarele trasaturi:
- mecanismul inducerii (humoral sau celular)
- dinamica desfasurarii in timp
- particularitatile manifestarilor patologice
- posibilitatea combaterii.
Gell si Coombs au definit 4 tipuri de reactii de hipersensibilitate:
- reactii de tip I: reactiile de anafilaxie (anafilaxia generalizata, reactiile de anafilaxie locala, denumite si stari atopice1 : astmul bronsic alergic, febra de fan, urticaria, reactia Arthus, maladia serului).
- reactii de tip II: reactii de citotoxicitate mediate de anticorpi
- reactii de tip III: reactiile de hipersensibilitate induse de complexele imune
- reactii de tip IV: reactiile de hipersensibilitate intarziata, mediate de limfocitele T(reactia la tuberculina, brucelina, lepromina etc; dermatitele de contact, reactia de respingere a grefei).
Reactiile de tip I, II si III sunt mediate de anticorpi, iar cele de tip IV sunt mediate de celule.
Dovezi pentru natura imunitara a reactiilor de hipersensibiltate:
- reactiile de hipersensibilitate necesita stimularea prealabila (sensibilizarea) organismului uman sau animal cu antigenul inductor (alergenul);
- intre momentul contactului cu doza sensibilizanta si momentul administrarii dozei declansatoare este necesara o perioada de timp (5-10 zile), pentru sinteza efectorilor reactiei (anticorpi) sau pentru expansiunea clonelor de limfocite. Dupa acest interval, organismul devine sensibil la declansarea starii de hipersensibilitate, numai la contactul cu acelasi alergen care a creat starea de sensibilizare sau cu un antigen inrudit, care da reactie serologica incrucisata cu alergenul inductor. Intre antigenul sensibilizant, cel declansator si starea de hipersensibilitate este o relatie specifica;
- organismele sensibilizate prezinta un raspuns imun de tip humoral sau celular. Starea de hipersensibilitate este mediata de efectorii raspunsului imun;
- reactiile de hipersensibilitate imediata se transfera prin ser de la un organism hipersensibil la unul sanatos. Pentru ca reactia de hipersensibilitate sa se manifeste, serul trebuie sa se injecteze intr-un tesut vascularizat. Reactiile de hipersensibilitate intarziata se transfera prin intermediul limfocitelor viabile.
Reactiile de hipersensibilitate imediata de tip I
Reactiile de hipersensibilitate de tip I, denumite si reactii anafilactice au cea mai mare frecventa si se manifesta foarte diferit, atat in ceea ce priveste intensitatea, cat si a organului tinta al reactivitatii.
|
Fig. 115 Reprezentare schematica a particularitatilor de evolutie a celor patru tipuri de reactii de hipersensibilitate. |
Antigenele care induc manifestarile reactiilor anafilactice se numesc alergene. Ele se gasesc in polenul unor plante, in praful de casa, in veninul insectelor sau in produse alimentare. Alergenele sunt un set de antigene, stimulatoare ale sintezei de IgE. Natura lor chimica este foarte heterogena. Un studiu german recent, releva ca organismul uman vine in contact cu circa 14000 de substante chimice: unele sunt substante alimentare, altele sunt ingerate odata cu alimentele, fiind adaugate in procesul industrial al prelucrarii. O categorie larga o formeaza substantele poluante. Din punct de vedere chimic, alergenele sunt glicoproteine si polizaharide de origine vegetala sau animala sau molecule mici, cu rol de haptene.
Haptenele sunt molecule organice sau anorganice, cu greutate mica, insuficienta pentru a fi antigene, dar devin alergene dupa cuplarea lor cu macromoleculele tisulare. De cele mai multe ori, haptenele alergice sunt substante de uz farmaceutic, de 500-1000 D. Dupa legarea covalenta ireversibila cu proteinele serice, rezulta un conjugat haptena-proteina, cu specificitate antigenica modificata si adeseori alergic. Toate medicamentele in stare nativa, dar si derivatii lor de degradare partiala pot sa se comporte ca haptene si sa devina alergene.
Intensitatea reactiilor de hipersensibilitate este dependenta de calea de patrundere in organism, de doza, de frecventa expunerii si de caracteristicile moleculare ale alergenului. Cel mai adesea, alergenele se clasifica in functie de calea de patrundere in organism. In raport cu calea de patrundere a alergenelor, se disting:
- alergene inhalate
- alergene ingerate
- alergene inoculate
Alergenele inhalate sunt glicoproteine din polen, din fungi, din praful animalelor de casa. Greutatea moleculara nu depaseste 50 kD, deoarece moleculele mai mari nu strabat membranele mucoase ale tractului respirator. Un alergen inhalat poate determina simptome respiratorii: rinita, astm. Crizele se produc numai in prezenta alergenului. Unele alergene sunt sezoniere (polen), altele sunt perene (praful de casa).
Polenul este, din punct de vedere cantitativ, cea mai importanta sursa de alergene. Plantele din familiile Gramineae, Compozitae, Betulaceae, Fagaceae produc polen alergic. Toate polenurile alergice provin de la plante polenizate de vant (anemofile). Ele produc cantitati mult mai mari de polen decat plantele entomofile si in perioada infloririi il elibereaza in atmosfera. Alergenele din polen sunt glicoproteine inrudite chimic si dau reactii incrucisate. Serul imun de iepure, fata de un alergen din polen precipita o diversitate de alergene polenice.
Sporii fungilor se gasesc in aer, in cantitate de circa 5 ori mai mare decat granulele de polen, dar produc mai putine stari alergice. In regiunile temperate, numarul sporilor fungici este maxim in timpul verii si scade in sezonul rece. Alergiile sunt produse de sporii a peste 80 de genuri de fungi: Alternaria, Penicillium, Aspergillus. Singura substanta alergica de origine fungica, izolata in stare pura, este penicilina.
Cantitatile de alergene care patrund pe cale respiratorie, sunt foarte mici: sub 1 μg/an.
Reactiile alergice la alergenele fungice apar la 30 de minute de la expunere, iar cele toxice, la 6-8 ore dupa ingestie. Reactiile toxice nu au substrat imunologic. Micotoxinele au greutate moleculara mica (sub 1 kD) si pot fi eliminate din extractele de alergene fungice, prin dializa.
Cele mai multe particule inhalate (mai mari de 10 μm), ca de exemplu, polenul si sporii mari sunt depozitate in nazofaringe si sunt asociate cu manifestari locale, nazale si/sau oculare, denumite generic "febra de fan". Particulele mai mici de 10 μm (dar in special cele mai mici de 5 μm) sunt antrenate cu curentul de aer inspirat, in caile inferioare, unde reactiile alergice tind sa se manifeste sub forma de astm. Sporii fungici difera ca dimensiuni si determina reactii alergice ale tractului respirator superior si inferior.
Alergenele ingerate se gasesc in compozitia unor alimente: in ou, ciocolata, capsuni, cirese, uleiul de ficat de peste, in semintele de Glycine max (soia), in faina unor cereale (grau, orz), in semintele de Arachys hypogea (alun de pamant) etc. La copii, laptele de vaca si soia sunt cauzele majore ale reactiilor alergice, urmate de cereale, oua si peste. Varsta manifestarilor este variabila: dupa primele zile de viata(fata de laptele de vaca), pana la doi ani. La 90% din cazuri, intoleranta dispare dupa trei ani. Cele mai comune manifestari sunt voma, diareea, colicile abdominale, ce apar intr-un interval variabil, de la cateva minute, pana la 1-2 ore dupa ingestie. Alergia la laptele de vaca si la peste este mai frecventa la populatiile care consuma cantitati mari ale acestor produse.
Alergenele alimentare traverseaza lumenul intestinal, la nivelul mucoaselor digestive. Bariera mucoasei gastrointestinale este expusa la un grup heterogen de antigene. Experientele cu molecule marcate cu peroxidaza de hrean sau cu feritina, au evidentiat ca unele antigene intacte si fragmente de antigene particulate din tubul digestiv, dobandesc accesul la tesuturile limfoide ale gazdei, in special la persoanele cu deficit al sintezei de IgA. Antigenele ce ajung in contact cu structurile limfoide ale mucoaselor, stimuleaza raspunsul imun al gazdei.
Penetrarea antigenelor la nivelul tractului digestiv se face pe doua cai:
- prin endocitoza de catre celulele intestinale absorbante, la polul luminal. Se formeaza fagosomi, in interiorul carora se produce digestia materialului endocitat, dar cantitati mici raman nedigerate si sunt exocitate in spatiul extracelular, la polul bazal. Starile de hipo- sau aclorhidrie favorizeaza tranzitul proteinelor prin mucoasa intestinala.
- patrunderea antigenelor alimentare la nivelul celulelor epiteliale M, ce acopera placile Peyer, foarte numeroase in regiunea distala a intestinului subtire. Aceste celule functioneaza ca adevarate "sonde de antigen", adica au capacitatea de a ingloba antigenele derivate in special din microorganisme si intr-o masura mai mica, antigene de origine alimentara.
Celulele M reprezinta un sistem de avertizare timpurie pentru sistemul imunitar al organismului. Ele sunt acoperite de un strat subtire de mucus, au microvili scurti, dar au capacitatea de a endocita antigene luminale, prin mecanismul pinocitozei. Celulele M sunt diferentiate, contin lizosomi si nu degradeaza antigenele pe care le pinociteaza, dar le transfera macrofagelor din foliculii subiacenti.
Desi au evoluat ca o modalitate strategica protectoare fata de antigenele luminale, totusi celulele M reprezinta poarta de intrare pentru microorganismele patogene, care la acest nivel isi dobandesc accesul la tesuturile subiacente mucoasei.
Studiile electrono-optice cu molecule marcate cu peroxidaza de hrean, au aratat ca acestea sunt transportate din lumenul intestinal si ajung in spatiul subiacent celulelor M, unde se gasesc limfocite si macrofage. Se stimuleaza astfel raspunsul imun local. Structurile limfoide asociate mucoaselor constituie prima bariera de protectie fata de antigenele tractului digestiv. Uneori, cantitatile relativ mari de antigene de origine bacteriana si alimentara, nu sunt anihilate local, de structurile limfoide ale mucoaselor si trec in mediul intern, ajungand la cea de a II-a bariera majora de protectie fata de antigene, care este ficatul. Circa 30% din numarul total de celule ale ficatului au capacitatea de a fagocita si epureaza sangele adus de vena porta din teritoriul digestiv.
Antigenele inoculate sunt proteine din veninul de insecte, in special Hymenoptere (albina, viespe), care contine 7-10 antigene. Alergenele din venin sunt diferite forme ale fosfolipazei A. Veninul de albina contine fosfataza acida, hialuronidaza, dopamina si norepinefrina si un peptid care produce degranularea mastocitelor.
Serurile imune obtinute pe diferite specii de animale (heteroantiseruri), dupa injectare la om, adeseori activeaza raspunsul imun al organismului receptor. Multi diabetici tratati cu insulina de origine animala sau cu insulina sintetizata in celule reprogramate prin metodele ingineriei genice, sintetizeaza anticorpi anti-insulina.
Agentii farmacologici, administrati in scop terapeutic sau diagnostic, pot cauza o varietate de dezordini imunitare, deoarece actioneaza ca haptene care se cupleaza cu diferite proteine tisulare, conferindu-le imunogenitate. Legarea covalenta a unui medicament ori a unui metabolit reactiv derivat prin metabolizarea lui, de o macromolecula, creeaza un conjugat haptena-macromolecula, inductoare a raspunsului imun specific. Legarea covalenta a celor doua molecule se numeste haptenare.
Se descriu doua tipuri de haptenare:
- haptenarea directa a celulelor (a moleculelor membranare) si a moleculelor extracelulare, sub actiunea compusilor chimici cu reactivitate nativa (intrinseca). De exemplu, penicilinele de semisinteza (benzil-penicilina, cefalosporinele), dar si alte medicamente se cupleaza cu diferite proteine serice, formand conjugate cu functie de alergene. Circa 10% din moleculele de penicilina injectata, se leaga covalent prin gruparea -NH2, de proteine plasmatice sau membranare. Celula poate lega mii de haptene β-lactamice, in cateva minute dupa tratament.
- haptenarea indirecta a moleculelor membranare sau libere, cu derivatii rezultati din catabolizarea partiala a unor molecule, care in stare nativa sunt putin reactive sau areactive.
Metabolizarea are loc in hepatocite, cheratinocite (si in alte celule) si poate crea intermediari reactivi ce formeaza legaturi covalente cu molecule carrier.
Uneori, catabolismul medicamentelor este concomitent cu sinteza proteica. Se produce haptenarea moleculelor in cursul sintezei (haptenare interna) si pot fi expuse ca antigene pe suprafata celulei.
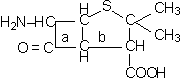
|
|
a = inel β-lactamic
b = inel tiazolidinic
Dupa fisiunea inelului β-lactamic rezulta gruparea peniciloil, principalul inductor al reactiilor anafilactice la om.
Metabolitii reactivi pot fi secretati in spatiul extracelular si se leaga cu proteine extracelulare.
Unii indivizi sunt predispusi la reactiile alergice fata de diverse medicamente, in special antiinfectioase. Daca un individ manifesta fenomene alergice fata de un compus antimicrobian, riscul alergiei fata de o alta clasa de compusi farmacologici creste de 9 ori. Circa 10% dintre adulti sunt alergici fata de o clasa de compusi si intra in categoria celor cu risc crescut fata de alti compusi farmacologici.
Reactiile de hipersensibilitate imediata de tip 1, induse de medicamente, se manifesta variat: anafilaxie, urticarie, angioedem.
Unele substante pot produce mai mult de un tip de reactie, la un organism sensibil. De exemplu, penicilina poate cauza o reactie anafilactica de tip I, o anemie hemolitica datorata reactiei citotoxice de tip II, dezordini functionale de tip III cu complexe imune sau o reactie de hipersensibilitate intarziata.
Reactiile de hipersensibilitate apar, de cele mai multe ori, in tesuturile bogate in mastocite: tegument, mucoase, mucoasa linguala, plaman, tractul gastrointestinal.
Daca reactia de hipersensibilitate imediata este localizata in mucoasa nazala si in conjunctiva oculara, simptomele includ rinoree, lacrimare, stranut, congestie nazala, cresterea numarului eozinofilelor in sange. Titrul IgE seric poate sa creasca sau sa ramana scazut.
Daca reactia de hipersensibilitate imediata este localizata in bronhii, manifestarea clinica este astmul alergic, caracterizat prin scurtarea si ingreunarea respiratiei.
Mediatorii reactiei de hipersensibilitate imediata de tip I
Reactiile de hipersensibilitate imediata de tip 1 se datoreaza sintezei unor izotipuri de anticorpi cu proprietati particulare.
Din punct de vedere functional, anticorpii sunt conventionali si citofili.
Anticorpii conventionali (Ig A, IgG, IgM) din ser se detecteaza in vitro, prin reactia de aglutinare, precipitare sau de fixare a complementului. In vivo, anticorpii conventionali se combina cu antigenul si il neutralizeaza.
Reactiile de hipersensibilitate de tip 1 se datoreaza sintezei anticorpilor citofili, cu proprietati functionale particulare.
Anticorpii citofili au proprietatea de a se fixa pe suprafata unor celule care au receptori pentru regiunea Fc si in primul rand pe mastocite, dar si pe bazofile, eozinofile, macrofage. Nu produc reactii secundare de aglutinare, precipitare sau de fixare a complementului.
Anticorpii citofili declansatori ai reactiilor de hipersensibilitate imediata se numesc reagine, deoarece produc modificari ale reactivitatii tisulare: maresc permeabilitatea vasculara prin intermediul histaminei, eliberata din mastocite. La om, reaginele majore sunt IgE si IgG4. IgE este termolabil (se inactiveaza la 56o, timp de 30 de minute), iar IgG4 este termostabil. Ambele izotipuri se sintetizeaza dupa imunizari naturale. IgE nu traverseaza bariera placentara. Timpul de injumatatire este de doua zile.
Sinteza lor se face dupa mecanismul clasic: dupa ce alergenul vine in contact cu mucoasele (respiratorie sau digestiva), penetreaza pelicula de mucus si celulele epiteliale, este fagocitat de macrofage, prelucrat si prezentat in asociatie cu moleculele CMH II, limfocitelor Th. Limfocitele Th sintetizeaza IL-2 care produce expansiunea clonala a limfocitelor B. Sub actiunea stimulatoare a IL-2, limfocitele B se transforma in plasmocite ce sintetizeaza IgE.
La persoanele normale, sinteza IgE este supresata de limfocitele Ts, iar concentratia de IgE seric variaza intre 17-450 ng/ml, adica 0,002% din totalul cantitatii de imunoglobuline serice. La indivizii atopici (cu predispozitie genetica pentru manifestarea reactiilor alergice), dupa contactul cu o doza mica de alergen, aproape totdeauna se sintetizeaza IgE, iar titrul seric creste de 1000 de ori. Nivelul seric crescut al IgE este o modalitate de diagnostic al starilor alergice, dar nivelul normal al IgE nu exclude starea alergica.
IgE se sintetizeaza local, in structurile limfoide, la poarta de intrare a antigenului, in plasmocitele din corionul mucoasei. Excesul de IgE trece in circulatie si se de mare afinitate ai bazofilelor, neutrofilelor, leaga pe receptorii pentru Fc ai celulelor endoteliului vascular, ai celulelor epiteliale alveolare pulmonare. Sangele transporta IgE la masocitele tisulare din tot organismul. IgE fixat pe celule reprezinta o proportie importanta din IgE total.
Desi IgE liber (din ser) are timpul de injumatatire de doua zile, IgE fixat pe suprafata celulelor este foarte stabil. Mastocitele raman sensibilizate luni de zile , datorita afinitatii foarte inalte a receptorilor lor pentru Fc al IgE. In stare legata, IgE este protejat de atacul proteazelor.
Celulele mediatoare ale reactiilor de hipersensibilitate imediata sunt in primul rand mastocitele si bazofilele .
Bazofilele sunt celule circulante , dar pot patrunde in focarul inflamator. Mastocitele sunt celule mononucleate si au doua localizari principale:
- in tesutul conjunctiv, in special in jurul vaselor sanguine din ficat, rinichi, splina;
- in mucoasele digestive si cea respiratorie.
In creierul uman sunt doua surse de histamina: mastocitele si neuronii histamiergici. Mastocitele se gasesc in zonele cele mai vascularizate ( eminenta mediana, glanda pineala, meninge ) si controleaza circulatia sangelui si permeabilitatea vaselor SNC. Neuronii histaminergici sunt limitati exclusiv la nivelul nucleului tuberomamilar din hipotalamusul posterior. Colateralele axonale se proiecteaza in toate zonele cortexului. Astfel, sistemul histaminergic central controleaza activitatea intregului creier.
Localizarea strategica a mastocitelor se coreleaza cu alterarile patologice care se produc la nivel vascular gastrointestinal, bronsic, cutanat, cerebral, ocular.
In citoplasma bazofilelor si mastocitelor, la microscopul electronic se observa granule electrono-dense, ce reprezinta pana la 40% din volumul celular. Ele contin mediatori preformati, cei mai importanti fiind aminele biogene (histamina si serotonina).
Mastocitele contin histamina (amina tisulara), detectata initial in tegument. In vivo, histamina se sintetizeaza prin decarboxilarea histidinei, reactie catalizata de histidin-decarboxilaza (o enzima citoplasmatica) si este depozitata in granule. O celula umana contine 2-3 pg de histamina.
Histamina este larg distribuita in tesuturile organismului uman. Histamina actioneaza asupra unei varietati largi de tipuri celulare: celule musculare netede, neuroni, celule endocrine, exocrine, celule sanguine, celulele sistemului imunitar. Efectele variate se produc prin receptori distincti: H1, H2, H3.
Serotonina (5-hidroxi-triptamina) se formeaza prin decarboxilarea moleculei de triptofan hidroxilat. Mastocitele umane nu contin serotonina. Serotonina s-a izolat initial din ser (tonina din ser), deoarece are efect vasoconstrictor. Produce edem si are rol important in reactiile anafilactice.
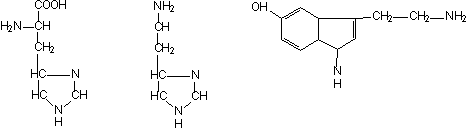
Histidina Histamina Serotonina (5-hidroxi-triptamina)
Mecanismul celular si molecular al reactiilor de hipersensibilitate imediata de tip I
Reactiile de hipersensibilitate imediata de tip 1 se desfasoara in mai multe stadii.
Faza I a este legarea anticorpilor citofili (IgE, care s-a sintetizat dupa contactul primar cu antigenul), de receptorii pentru Fc ε ai mastocitelor locale. Alti factori activatori ai mastocitelor sunt:
- alergenul specific sau unul inrudit cu cel inductor al sintezei IgE;
- diferite lectine
- anticorpi anti-IgE, anticorpi anti-idiotipici, anticorpi anti-receptor Fc ε;
|
Fig. 116 Activarea masto-citelor este mediata de legarea incrucisata a recep-torului Fc ε. Aceasta poate fi realizata de legarea antigenului de IgE fixat pe receptorii pentru Fc ε, de anticorpii bivalenti care recunosc determinantii izo-tipici ai regiunii Fc a IgE, de anticorpii anti-idiotipici ai regiunii Fab ai IgE, de anticorpii anti-receptor care se fixeaza direct pe recep-torul pentru Fc ε, de dimerii bivalenti de IgE obtinuti cu agenti chimici de polime-rizare sau de lectinele care se leaga de resturile glu-cidice ale IgE. Antigenele si anticorpii monovalenti nu activeaza mastocitele, deoa-rece nu realizeaza legarea incrucisata a receptorilor (dupa Roitt, 1993). |
- neuropeptide endogene, fapt care explica alergia determinata de stimularea sistemului nervos
- anafilatoxinele C3a si C5a
- peptidele bacteriene ce contin formil-metionina
- agenti fizici (de exemplu, temperatura scazuta)
- diferite medicamente.
In faza a II-a se produce activarea mastocitelor si degranularea lor.
Alergenul declansator trebuie sa fie multivalent pentru a lega incrucisat doua molecule de IgE fixate pe celula efectoare. Dupa legarea incrucisata a receptorilor pentru Fc ε, membrana granulelor fuzioneaza cu membrana citoplasmatica si elibereaza continutul.
|
Fig. 117 Alti stimuli activatori ai mastocitelor. Anafilatoxinele C3a, C5a sau substantele medica-mentoase (ionoforii de Ca2+, codeina, morfina, ACTH sintetic) activeaza mastocitele pe o cale directa. Toate aceste substante induc patrun-derea Ca2+ in mastocit, declansatoare a fenome-nelor biochimice care duc la degranularea si eliberarea mediatorilor. |
![]()
Eliberarea mediatorilor preformati
- histamina
- triptaza - activeaza C3
- heparina
Moleculele de IgE pot fi legate incrucisat de lectine (PHA, Con A), prin asocierea lor cu resturile glucidice ale regiunii Fc. Astfel se explica urticaria produsa de capsuni si cirese, care contin cantitati mari de lectine. Stimulul activator al mastocitelor este transmis prin receptorii pentru IgE. Activarea se datoreaza legarii incrucisate a doua situsuri Fab, printr-o molecula de alergen. , care leaga IgG cu afinitate mult maigMastocitele au si receptori pentru Fc mica. Degranularea mastocitelor este precedata de influxul masiv de Ca2+. Granulele pline cu mediatori preformati, migreaza la periferia celulei, fuzioneaza cu membrana externa si elibereaza continutul: histamina, proteaze neutre(triptaza, chimaza, carboxipeptidaza), proteoglicani (heparina, condroitin-sulfatul).
|
Fig. 118 Inducerea si mecanismele efectoare ale hipersensibilitatii de tip I. Prelucrarea antigenului se face la nivelul mucoaselor. Reexpunerea la alergen declanseaza degranularea mastocitelor si producerea mediatorilor ce determina simptome alergice (dupa Mirakian, 1998). |
Triptaza (prezenta in mastocite, dar absenta in bazofile) produce constrictia muschilor netezi ai bronhiilor, iar chimaza stimuleaza secretia mucoasei bronsice.
Proteoglicanii din granulele mastocitelor au rol in impachetarea mediatorilor preformati in granule.
Heparina este proteoglicanul predominant in mastocitele pulmonare umane. Mastocitul este singura sursa endogena de heparina la om si la rozatoare. Heparina are efect anticoagulant.
Mastocitele contin SOD si peroxidaza.
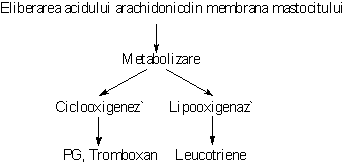
Degranularea activeaza o lipaza care mobilizeaza acidul arachidonic din membrana mastocitului. Prin metabolizarea acestuia se sintetizeaza mediatori noi ai reactiei de hipersensibilitate imediata. Astfel, din acidul arachidonic, pe calea lipooxigenazei se sintetizeaza leucotriene, iar pe calea ciclooxigenazei se formeaza prostaglandine si tromboxan.
Efectele mediatorilor mastocitari
Manifestarile reactiilor alergice, inclusiv alergia respiratorie, pot sa se produca in doua faze:
- faza timpurie, care se desfasoara in cateva minute, datorita eliberarii mediatorilor preformati;
- faza tardiva se produce la 3-4 ore dupa expunerea la alergen, datorita infiltratului celular, ca raspuns la mediatorii fazei timpurii.
In prima etapa, actiunea histaminei consta in contractia muschilor netezi ai tractului respirator si digestiv. Histamina produce constrictia celulelor endoteliale si creste permeabilitatea vaselor mici. Acelasi efect il are serotonina. Prin constrictia celulelor endoteliale si cresterea permeabilitatii capilare, se produce edemul tisular.
In etapa a II-a, histamina produce dilatarea vaselor periferice si rezultatul este scaderea brutala a tensiunii arteriale. Socul hipotensiv este una din manifestarile dramatice ale anafilaxiei generalizate.
Alti mediatori ai fazei a II-a sunt chininele. Ele se formeaza din chininogenul plasmatic si sunt polipeptide mici:
- metionilchinina (Met-
- bradichinina(Arg-(Pro)2-Gly-Phe-Ser-Pro-Phe-Arg)
- calidina (lisil-bradichinina):
Chininogenul este alcatuit din 11 aminoacizi si (acid sialic)n, la una sau la ambele extremitati. Este o α-globulina (2 mg/ml ser).
Chininele au efect vasodilatator si maresc permeabilitatea capilara, producand edem.
In faza a III-a se sintetizeaza alti mediatori, care intretin in timp diferitele efecte initiate de mediatorii eliberati din mastocite, prin efectul chimiotactic fata de celulele sanguine:
ECF (eozinofil-chemotactic factor) determina afluxul de eozinofile
NCF (neutrofil-chemotactic factor)
PAF (factorul activator al plachetelor).
Leucotrienele (LC4, LD4, LB4, LE4) determina contractia de durata a muschilor netezi, edem al mucoaselor si stimularea secretiei lor.
Exemple de reactii de hipersensibilitate de tip I
Prima reactie de hipersensibilitate imediata a fost descrisa de Prausnitz si Kustner.
Reactiile de hipersensibilitate imediata pot sa se produca la orice organism al unei specii si se numesc reactii anafilactice sau se manifesta numai la anumiti indivizi predispusi genetic si se numesc atopii (stari atopice).
Reactiile anafilactice (ana = opus; phylaxis = protectie) se caracterizeaza printr-o dinamica exploziva a manifestarilor, in decurs de 3-4 minute. Clinic, termenul de anafilaxie semnifica sindromul care rezulta din eliberarea unor mediatori preformati si generati de novo in mastocite, care determina o stare opusa celei de protectie. Manifestarile clinice ale reactiilor de hipersensibilitate sunt dependente de calea de patrundere a alergenului si de cantitatea de histamina eliberata. Reactiile cutanate medii, cu eritem, urticarie, prurit, dureri de cap, sunt asociate cu un nivel scazut al histaminei plasmatice (sub 1 ng/ml).
Daca alergenul este injectat intravenos, raspunsul este sistemic, adica are loc o anafilaxie generalizata, care poate duce la soc vascular hipotensiv si asfixie secundara prin constrictie bronsica si laringiana. Daca sfarsitul nu este letal, recuperarea functionala se poate face intr-o ora. Socul vascular se datoreaza iesirii plasmei in spatiul extravascular, rezultand scaderea volumului sanguin. Scaderea debitului cardiac duce la hipoxie si acidifierea mediului intern, antrenand si insuficienta respiratorie.
Daca alergenul este injectat in piele, reactia este limitata la locul injectiei si se numeste anafilaxie cutanata, insotita de eritem (urticarie), angioedem, edem laringian, bronhoconstrictie, hipotensiune, aritmie cardiaca.
Anafilaxia generalizata (sistemica) este forma cea mai grava a hipersensibilitatii imediate. Mult timp s-a considerat ca anafilaxia generalizata este o manifestare patologica, indusa numai in conditii experimentale la animalele de laborator. In realitate, atat la om cat si la animale se produc ambele tipuri de reactii anafilactice, atat generalizate, cat si locale.
Prima reactie de anafilaxie generalizata a fost descrisa la caine. In 1902, Richt si Portier studiau biologia meduzelor si toxicitatea extractelor apoase si glicerinate din celenterate, asupra mamiferelor. Administrate in doze mari, extractele produc moartea imediata a cainelui, datorita socului toxic, dar dozele mici sunt suportate. O injectie secundara la cateva saptamani mai tarziu, produce o reactie anafilactica generalizata, cu paralizia muschilor respiratori si socul fatal. Instalarea starii de soc este accelerata de injectarea intravenoasa a dozei secundare. La autopsie se observa edemul mucoasei intestinale, a tesutului pulmonar, congestia ficatului.
La cobai, anafilaxia se induce prin injectarea intravenoasa a unei doze mici (1mg) de albumina serica bovina. La 8 zile dupa injectarea dozei sensibilizante, animalul devine sensibil la socul anafilactic si are sensibilitate maxima la 21 de zile. Doza declansatoare a socului este mai mare. Raspunsul anafilactic este prompt si manifestarile incep in primele zeci de secunde: piloerectie dorsala, agitatie motorie, stranut, tuse zgomotoasa, cianozarea mucoaselor, pierderea reflexelor de mictiune si defecatie, blocaj respirator, convulsie, moarte. Tabloul acestor modificari patologice se succede in cateva minute. Muschii netezi bronsici ai cobaiului sunt foarte sensibili la histamina si bronhoconstrictia fatala este caracteristica anafilaxiei la aceasta specie.
Reactiile anafilactice generalizate la om se manifesta in primul rand fata de alimente si se exteriorizeaza prin urticarie generalizata. Urticaria afecteaza 15-23% din populatie Suportul manifestarilor patologice care insotesc aceste reactii, este tegumentul.
Anafilaxia generalizata la om, este declansata si de alti factori: penicilina, aloseruri, heteroseruri, veninul insectelor (albina, viespe) si se manifesta prin urticarie generalizata, spasmul muschilor bronsici, edem laringian, dispnee severa, cianoza, soc hipotensiv. Intensitatea manifestarilor este dependenta de gradul de sensibilitate. Sensibilitatea alergica a pacientilor variaza in limita a circa 1/1000.
Raspunsul sistemic cu reactii tegumentare generalizate, tulburari gastrointestinale, tahicardie, aritmie cardiaca, hipotensiune medie, este asociat cu cresterea concentratiei plasmatice a histaminei peste nivelul de baza. Reactia ampla care pericliteaza viata, cu hipotensiune severa, fibrilatie ventriculara, spasmul muschilor bronsici, stop cardiac si respirator, este asociata cu nivelul crescut al histaminei, de 12 ng/ml. La om, histamina are efect vasodilatator asupra vaselor periferice, cu scaderea presiunii vasculare.
Hipersensibilitatea imediata la penicilina produce circa 500 decese/an/glob. Reactia este declansata de penicilina nativa sau de derivatii rezultati din degradarea ei (acidul peniciloic, acidul penicilenic).
Anticorpii
anti-penicilina apartin izotipurilor IgE si IgG4 si se
gasesc chiar in serul persoanelor netratate cu penicilina.
Probabil ca sinteza lor a
fost indusa de penicilina din carne. Originea penicilinei este in masa
miceliana de la fabricile de antibiotice, care se adauga in hrana
animalelor, ca stimulator al cresterii.
Alergenele inhalate produc manifestari locale, la poarta de intrare.
Reactii atopice. Termenul de atopie defineste starile de hipersensibilitate imediata cu substrat ereditar, limitate la om. Starile atopice se manifesta la 10-20% din populatie, sub forme variate.
Reactiile atopice au un tablou clinic mai simplu si se produc atunci cand reactia alergenului cu anticorpii specifici are loc pe suprafata unei mucoase sau in epiteliul tegumentar.
Rinita alergica sau febra de fan este forma cea mai simpla a atopiei si se manifesta prin secretie abundenta a glandelor epiteliului nazal, iar astmul bronsic, prin contractia muschilor cailor respiratorii. Cele doua forme extreme de manifestari alergice sunt determinate de alergenele inhalate, care se gasesc in polen, in praful de casa(ce contine alergene de acarieni), in parul animalelor. Foarte raspandite sunt alergiile la polenul de Ambrozia, o planta care raspandeste o cantitate mare de polen (100 kg/planta/an). Grauncioarele de polen vin in contact cu mucoasa nazala si respiratorie, cu conjunctiva oculara si determina o stare de sensibilizare a aparatului respirator. Reexpunerea la polen in sezonul urmator, declanseaza reactia alergica, cu urmatoarele manifestari: eritemul conjunctivei oculare, edemul mucoasei nazale si secretia apoasa abundenta, stranut.
Intensitatea simptomelor depinde de nivelul cantitativ al expunerii la polen, dar si de reactivitatea organismului. Exista situatii grave, inexplicabile, cu crize ample, dupa o expunere minimala.
Daca rinita are caracter sezonier sau este dependenta de conditiile de mediu, aproape sigur are caracter alergic. Rinita alergica perena se datoreaza contactului cu praful de casa si de animale.
Mecanismul rinitei alergice este diferit de al altor manifestari alergice. In primul rand, exista diferente functionale intre mastocitele mucoaselor si ale tesutului conjunctiv. Mastocitele mucoasei nazale(denumite MCT) contin histamina si triptaza, iar dupa stimularea alergenica, se degranuleaza, dar concentratia plasmatica a histaminei nu se modifica. Mediatorii mastocitari produc o hiperemie a mucoasei nazale, edem si transudarea lichida. Inflamatia nazala se datoreaza transudarii, in care albumina este un marker mult mai stabil decat mediatorii din categoria citochinelor.
|
Fig. 119. Mecanismul rinitei alergice (dupa Frankland, 1998). |
In al II-lea rand, manifestarile rinitei sunt dependente de particularitatile unice ale vascularizatiei nazale, conferite de tesutul venos erectil. Variatiile volumului sanguin din tesutul venos erectil regleaza rezistenta tractului nazal la coloana de aer. Umplerea cu sange a acestui tesut produce congestia mucoasei. Curgerea sangelui este controlata de fibrele nervoase vegetative. Inervatia simpatica este vasoconstrictoare si diminua fluxul sanguin, iar tonusul simpatic are activitate ciclica. De aceea, mucoasa nazala are o perioada de repaus la fiecare 2-4 ore, iar caile nazale se pot obtura alternativ.
Faza primara a rinitei (stranut, rinoree, obstructie nazala) este dependenta de histamina din mastocite, de leucotriene si prostaglandine, iar faza tardiva (obstructie nazala ampla prin hiperreactivitate si anosmie) este dependenta de bazofile.
Astmul alergic, o varianta complexa a reactiilor atopice, este o maladie cronica inflamatorie a plamanului, cu o prevalenta crescanda a morbiditatii si mortalitatii in ultimele doua decenii. Desi s-a considerat ca este o boala a muschilor netezi ai cailor respiratorii, in ultima decada s-a acceptat ca astmul este, in primul rand, o boala inflamatorie. Structural, caile aeriene ale astmaticilor se caracterizeaza prin inflamatie cronica, cu infiltrarea intensa a mucoasei bronsice, cu limfocite, eozinofile si mastocite, cu descuamarea epiteliului, hiperplazia celulelor mucoase, ingrosarea submucoasei. Aceste modificari se asociaza cu manifestarile clinice, care includ obstructia cailor aeriene si hiperreactivitatea cailor aeriene.
Astmul bronsic alergic are determinism multifactorial, atopia (predispozitia genetica de a sintetiza IgE ca raspuns la aeroalergenele comune) fiind cel mai comun factor predispozant. Dupa expuneri repetate la doze mici de alergene, indivizii atopici sintetizeaza IgE. Expunerea secundara la alergene initiaza raspunsul imun humoral. Evenimentele reactiei alergice, consecutive expunerii la alergen, se desfasoara in doua faze: faza timpurie, de ordinul minutelor, consta in raspunsul bronhospastic si faza tardiva, de ordinul orelor, ce consta in raspunsul inflamator.
Raspunsul rapid se caracterizeaza prin edemul mucoasei, cresterea tonusului muschilor netezi si ingustarea cailor respiratorii, asociata cu degranularea mastocitelor. Manifestarile astmatice imediate sunt dependente de eliberarea histaminei din mastocite. Pacientii cu astm alergic au o concentratie de IgE de 6 ori mai mare decat astmaticii nealergici, ceea ce confirma relatia dintre concentratia IgE si starea patologica.
Unii astmatici alergici dezvolta raspunsul de faza tardiva, la 3-6 ore dupa stimularea cu alergenul, care, in absenta terapiei, poate sa persiste cateva zile. Raspunsul tardiv, este asociat cu migrarea eozinofilelor, dar si a neutrofilelor si limfocitelor din sange, in parenchimul pulmonar. Limfocitele TCD4 au rol esential in patogeneza astmului. In plamanul astmatic, alergenul este inglobat, prelucrat si prezentat de macrofagul alveolar, de celulele epiteliale ale mucoasei si de celulele dendritice. Limfocitele Th-2 secreta IL-4, 5, 6, 9, 10, 13, stimulatoare ale sintezei de IgE.
Alergiile la alimente formeaza o categorie de stari atopice foarte comune la copii si se manifesta fata de alergene din lapte, oua, ulei de peste, ciocolata, capsuni, cirese, portocale. Manifestarile reactiilor alergice la alimente pot fi locale sau generalizate.
Reactiile alergice locale se caracterizeaza prin eruptii urticariene (pustule eritematoase) la nivelul mucoasei bucale, insotite uneori de tulburari digestive (colici abdominale, diaree). La varsta adulta, subiectii vor manifesta reactii alergice respiratorii.
Nu toate urticariile tegumentare sunt determinate de reactii alergice. Urticaria neimuna de contact poate fi declansata de agenti chimici (salicilati, sulfiti, acidul benzoic, acidul sorbic, acidul nicotinic si esterii sai din alimente, din guma de mestecat, din sampoane, din parfum, din unguente) sau de agenti fizici (traumatisme, frigul). Acestia stimuleaza eliberarea factorilor chimici inductori ai urticariei. Mecanismele ei nu se cunosc, dar efectele se manifesta asupra endoteliului vascular. Simptomele sunt arsura, usturimea, pruritul, edemul si eritemul.
Urticaria este acuta sau cronica. Cea cronica se manifesta prin aparitia zilnica sau aproape zilnica a leziunilor tegumentare, pentru un interval de cel putin 6 saptamani, cauzata de stimuli fizici.
In toate cazurile de urticarie, imuna sau neimuna, pe tegument apar leziuni eritematoase (macule) si edematoase, insotite de prurit. Urticaria implica dermul superficial.
Leziunea cutanata eruptiva este datorata cresterii permeabilitatii capilarelor si dilatarii venulelor. Plasma paraseste patul vascular si determina formarea pustulei tegumentare pruriginoase, inconjurata de o zona de culoare rosie intensa. Lichidul veziculelor eruptive este infiltrat cu limfocite si PMNN. Eruptia tegumentara dispare la 24-48 ore, fara urme in cazul reactiilor acute sau persista mai multe zile la cele cronice.
In manifestarile cu angioedem, edemul se extinde subcutan sau submucos si leziunile pustulare sunt mai mari si pot sa dureze 72 de ore. Se manifesta la nivelul buzelor, limbii, pleoapelor, dar poate fi afectata orice zona corporala.
Pseudoalergii. Simptomele clinice ale alergiilor (urticarie, rinita, astm, socul anafilactic) apar adeseori din cauze nealergice. Numarul lor depaseste pe acela al alergiilor. Iata principalele cauze:
- histamina se gaseste in cantitati mari in unele alimente si activeaza mediatorii specifici ai reactiilor alergice;
- histidina din alimente este precursorul histaminei;
- tiramina de origine alimentara determina dureri de cap pulsatile si hipertensiune;
- feniletilamina produce migrena;
- benzoatul de origine alimentara si din acidul benzoic produce astm si rinita;
- bisulfitul de sodiu, bioxidul de sulf si metabisulfitii (conservanti) produc simptome ale tractului gastrointestinal, distrug vitaminele B, produc dureri de cap, astm;
- glutamatul monosodic (agent de aromatizare) produce astm;
- colorantii alimentari (in special culoarea galbena) produc astm si alte reactii alergice;
- aspirina induce astm, urticarie, angioedem (Girard, 1998).
Testarea starilor atopice
Pentru prevenirea accidentelor provocate de reactiile de hipersensibilitate imediata, in clinica, inainte de administrarea unor medicamente, se testeaza eventualele manifestari de hipersensibilitate.
O problema esentiala pentru diagnosticul si tratamentul alergiilor, este gradul inalt de variabilitate biochimica a alergenelor, de la un lot la altul. Preparatele de alergene sunt foarte heterogene, iar cantitatea de alergen specific, din cele mai multe extracte comerciale, variaza de la cateva procente pana la 50% din proteina totala. Extractele sunt amestecuri complexe, cu cantitati variabile de alergene majore, alergene minore si proteine irelevante. Extractele fungice contin proteine, glucide, enzime proteolitice, enzime glicolitice, in cantitati variabile. De aceea, efortul major s-a orientat in directia clonarii alergenelor de importanta clinica majora. Majoritatea alergenelor s-a clonat si unele sunt disponibile la un nivel moderat sau chiar inalt de puritate. Disponibilitatea preparatelor antigenice omogene, cu puritate si activitate biologica definite, va avea un impact major asupra diagnosticului si probabil, asupra terapiei dezordinilor alergice.
Multe alergene, in special cele de natura proteica, se prezinta intr-o multitudine de izoforme naturale. Ele sunt rezultatul variatiei alelice a numeroaselor proteine vegetale si fungice, foarte asemanatoare ca secventa a aminoacizilor, dar au proprietati distincte de combinare cu anticorpii sau de activare a clonelor de limfocite T specifice.
Existenta numeroaselor izoforme ale aceleiasi molecule de alergen, este probabil, asociata cu grade diferite de recunoastere a epitopilor sai, de catre limfocitelor T si B si, ca o consecinta, cu grade diferite de sensibilitate. Ca dovada, anticorpii de la diferiti pacienti alergici, leaga numai unele izoforme ale alergenului.
Preparatele de alergene pentru diagnostic sunt heterogene si deriva din surse naturale care contin majoritatea sau toate izoformele de alergen. Inlocuirea acestor alergene cu cele clonate, pentru diagnostic, va rezolva unele probleme, dar va crea altele. Unii cercetatori apreciaza ca majoritatea indivizilor raspund la un numar limitat de epitopi ai oricarui alergen. De aceea se considera ca un numar mic de alergene (circa 40) ar putea sa fie suficiente pentru a identifica majoritatea indivizilor alergici.
Testele cutanate constau in injectarea subcutana a alergenelor la diferite dilutii. La organismul alergic, reactia de hipersensibilitate se manifesta in cateva minute, prin aparitia la locul injectarii, a unei reactii inflamatorii pruriginoase, inconjurata de o zona mai extinsa de eritem. Reactia de hipersensibilitate la proteinele salivei de tantar are acelasi aspect. Alergenul injectat (sau inoculat) se leaga specific de IgE fixat pe mastocitele din piele. Mastocitele elibereaza histamina in cateva minute, cauzand edemul localizat, eritem (vasodilatatie) si prurit. Leziunile locale ale reactiei de hipersensibilitate imediata se reproduc prin injectarea unei cantitati mici de histamina.
Determinarea cantitativa a IgE seric al pacientului, ramane cel mai bun criteriu de diagnostic. IgE se dozeaza prin metoda RIA:
- alergenul (de exemplu, penicilina) se fixeaza pe suportul inert (dextran);
- proba de ser de cercetat se pune in contact cu alergenul fixat pe suport. IgE din ser se fixeaza pe alergenul imobilizat;
- se adauga ser imun de iepure, care contine IgG marcat radioactiv, anti-IgE uman;
- se masoara indirect cantitatea de IgE fixata pe suport, prin masurarea cantitatii de IgG radioactiv legat specific.
Combaterea starilor de hipersensibilitate imediata de tip I se face prin evitarea alergenelor, prin terapie medicamentoasa si prin imunoterapie.
Masurile de evitare a alergenelor sunt ideale pentru prevenirea reactiilor: evitarea alimentelor, medicamentelor, controlul prafului de casa, evitarea contactului cu animalele al caror praf este alergic, combaterea mucegaiului din incaperile locuite.
Medicamentele antihistaminice au efect antagonic fata de histamina, deoarece intra in competitie cu receptorii pentru histamina ai celulelor tinta. Astfel, se poate bloca cresterea permeabilitatii vasculare, vasodilatatia, contractia muschilor bronhiilor si ai mucoasei gastrointestinale. Rinita alergica necesita terapie antihistaminica locala. Cromoglicatul de sodiu protejeaza pacientii astmatici. Teofilina(o metilxantina) relaxeaza muschii netezi bronsici.
Imunoterapia sau hiposensibilizarea consta in administrarea planificata a alergenului la un pacient pentru a diminua sinteza IgE. Rinita alergica, astmul bronsic si anafilaxia la veninul de insecte pot raspunde la aceasta terapie. Alergenul este injectat saptamanal, in doze gradat crescande. Dupa o crestere initiala a IgE circulant, se produce declinul. Locul IgE este luat de IgG care fixeaza alergenul si-l impiedica sa stimuleze mastocitele, blocand reactia alergica.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2454
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2026 . All rights reserved